
HERICHTHYS MINCKLEYI – KORNFIELD & TAYLOR 1983
Considéré comme synonyme de Cichlasoma pendant plusieurs décennies, le genre Herichthys compte actuellement une dizaine d’espèces, toutes originaires d’Amérique centrale.
Parmi toutes ces espèces dont certaines sont très connues et prisées chez les aquariophiles, il en existe une qui vient d’un milieu exceptionnel et qui l’est tout autant : Herichthys minckleyi.

Parmi les vingt et une espèces de poissons qui peuplent la vallée, c’est « Herichthys minckleyi » qui a suscité le plus d’intérêt, en raison de son histoire évolutive fascinante.
Ce cichlidé est considéré comme une espèce polymorphe présentant trois morphotypes différents, qui cohabitent plus ou moins entre individus de la même espèce dans les cours d’eau de la vallée.

Selon la taxonomie traditionnelle, en 1974, des scientifiques étudiant la faune piscicole de la vallée ont suggéré que l’ampleur des différences observées entre les différents morphotypes de ce cichlidé justifiait amplement la création d’au moins trois espèces distinctes.
Espèce unique, endémique et menacée, le cichlidé de Minckley a attiré l’attention de chercheurs du monde entier, désireux de comprendre le polymorphisme trophique.

De plus, ce poisson contribue de manière précieuse à la biodiversité d’une région du monde incroyablement unique et exceptionnelle.

Plus d’informations dans ce qui suit …

–
REPARTITION
Herichthys minckleyi a été répertorié en 1983 pour la première fois par les chercheurs Irv KORNFIELD et Jeffery TAYLOR, le cichlidé de Minckley est endémique du bassin de Cuatro Ciénegas situé à Coahuila, au Mexique.

Les points indiquent les sites échantillonnés et les lettres la distribution actuelle de la famille des Cichlidae au nord de Punta Del Morro : (A) Herichthys minckleyi ; (B et C) Herichthys cyanoguttatus ; (D) Herichthys cyanoguttatus San Fernando ; (E) Herichthys cyanoguttatus San Ignacio Baths ; (F) Herichthys teporatus ; (G et H) Herichthys carpintis et H. pantostictus ; (I) Herichthys carpintis et Herichthys pratinus ; (J) Herichthys tamasopoensis, Herichthys pame et Herichthys steindachneri ; (K) Herichthys labridens et Herichthys bartoni ; (L) Herichthys molango.
Cette autre carte montre les bassins versants et l’hydrologie de surface des Herichthys.

–
A PROPOS DE CUATRO CIENEGAS
Parmi la faune de Cuatro Ciénegas, il existe un cichlidé endémique à cette seule région de Cuatro Ciénegas : « Herichthys minckleyi » : La répartition géographique de la « mojarra de Cuatro Ciénegas » Herichthys minckleyi, décrit pour la première fois par les chercheurs Irv KORNFIELD et Jeffery TAYLOR en 1983, est restreinte uniquement à Cuatro Ciénegas.
Herichthys minckleyi est aussi l’espèce la plus nordique du grand groupe « Herichthys cyanoguttatus ».
D’autre part, Herichthys minckleyi qui est l’un des poissons les plus courants du bassin est, de surcroit, un exemple classique de polymorphisme trophique.
Dans ce bassin inter-montagneux, ce cichlidé « Herichthys minckleyi » se rencontre dans les sources, les ruisseaux, les étangs et les lagunes et c’est l’espèce la plus septentrionale du groupe Herichthys cyanoguttatus.
Le bassin de Cuatro Ciénegas est particulièrement réputé pour sa faune piscicole diversifiée, et Herichthys minckleyi y apporte une contribution remarquable.
En effet, Herichthys minckleyi est l’un des poissons les plus communs de la vallée de Cuatro Ciénegas et, par conséquent, il constitue donc un maillon important du réseau alimentaire de cette vallée.

Carte montrant la région de Cuatro Ciénegas dans l’État mexicain de Coahuila.

Carte localisant la région de Cuatro Ciénegas dans l’État mexicain de Coahuila.

Carte localisant la région de Cuatro Ciénegas dans l’État mexicain de Coahuila.

Carte de répartition des Herichthys minckleyi dans Cuatro Ciénegas.
Cette espèce de cichlidé partage cet habitat avec la « mojarra del Norte », Herichthys cyanoguttatus avec un espace de distribution ouvert situé à 800 kilomètres du Tropique du Cancer au Rio Diablo au Texas, États-Unis.

« Mojarra del Norte », Herichthys cyanoguttatus
Du point de vue climatique et géographiquement, il est situé dans le désert de Chihuahua ou sur le plateau nord, dans la région centrale de l’État de Coahuila.
Le bassin de Cuatro Ciénegas est un bassin fermé intermontagneux.
La vallée de Cuatro Ciénegas est un écosystème désertique unique avec des niveaux d’endémisme exceptionnellement élevés.
Dans le nord du Mexique, dans l’État de Coahuila, la vallée de Cuatro Ciénegas, il est possible de découvrir ce lieu unique au monde qui est l’un des sites les plus biodiversifiés au monde où certains des organismes présents ici, encore actifs, sont les mêmes qui ont vu le jour il y a environ 3 500 millions d’années, et sont les premiers responsables de la production d’oxygène dans notre atmosphère.
C’est aussi le deuxième plus grand désert d’Amérique du Nord et la plus grande éco-région du Mexique.
Le climat y est aride, avec une pluviométrie moyenne inférieure à 200 mm.
Les températures estivales diurnes dépassent parfois 44°C et températures hivernales pouvant chuter en dessous de 0°C (Minckley, 1969).


GÉOGRAPHIE DE CUATRO CIÉNEGAS La vallée de Cuatro Ciénegas, conformément à la forme de W (rouge), se trouve en le désert Chihuahuense du nord du Mexique (A). Herichthys cyanoguttatus a une distribution géographique beaucoup plus grande (bleu) que Herichthys minckleyi (rouge), qui se trouve uniquement dans Cuatro Ciénegas. La vallée de Cuatro Ciénegas contient de nombreuses pozas alimentés par des sources et d’autres habitats aquatiques (B). Les lieux d’étude courants sont : Churince, Juan Santos, Tierra Blanca, Mojarral Oeste, Mojarral Este, Escobedo, Los Remojos, Candido Los Gatos.
Malgré le climat aride, le bassin abrite un vaste système de sources, ruisseaux et bassins.
–
De même que les îles Galapagos de l’Équateur, la plupart de ces habitats aquatiques sont géographiquement séparés et constituent des laboratoires naturels indépendants pour l’étude des processus écologiques et évolutifs (HULSEY & Al., 2016).
On estime que sur les 184 000 hectares la surface de la réserve qui possède plus de 400 bassins, de tailles, profondeurs, composition minérale de l’eau et températures variées, chacun hébergeant des formes de vie uniques.
Le Cuatro Ciénegas est un écosystème désertique unique avec des niveaux d’endémisme exceptionnellement élevés, c’est-à-dire qu’il abrite de nombreuses espèces végétales et animales que l’on ne trouve nulle part ailleurs dans le monde.
Bien qu’il y ait probablement eu des liens avec le río Grande dans le passé géologique, la région se caractérise aujourd’hui par son isolement.

Ces sources varient en taille, allant de quelques mètres carrés à plusieurs hectares.
Ce sont souvent des sources chaudes avec des températures pouvant atteindre environ 34 °C.
Mais les sources froides prennent parfois leur source à quelques mètres des sources chaudes et se rejoignent dans des ruisseaux, des rivières et des marais.
La vallée est entourée de crêtes montagneuses.
Dans la vallée, les eaux sont moins séparées qu’il n’y paraît.
De nombreux plans d’eau sont reliés entre eux par le moyen.
Les rivières souterraines et, de plus, les fortes pluies peuvent créer des connexions temporaires dans de nombreux endroits.
C’est MINCKLEY, un entomologiste de l’Université de Rochester, spécialisé sur le désert, qui a décrit de nombreux aspects de la vallée, sa géologie et sa biote aquatique et ce poisson fantastique vivant à cet endroit.
Dans le centre-nord du Coahuila, au Mexique, il y a cette vallée de 700km2 se situant à environ 740 mètres d’altitude et abritant au minimum plusieurs centaines de bassins à source allant de quelques mètres carrés à plusieurs hectares.
En cet endroit, la température de l’eau varie fortement entre les bassins et les sources chaudes et froides émergent souvent à quelques mètres les unes des autres, se fondant en ruisseaux, rivières et marais.

Riche d’écosystèmes nourris par des sources pauvres en azote et en phosphore, ce bassin présente étonnement un niveau élevé d’endémisme.
En fait, les niveaux d’endémisme y sont très élevés tant dans les communautés terrestres que celles aquatiques Mais aussi planent des menaces imminentes sous forme de déshydratation et d’espèces exotiques, autant d’éléments qui font de la conservation un enjeu important dans ce qui est aujourd’hui une zone naturelle protégée fédérale pour la flore et la faune.
Le bassin reçoit peu (<200 millimètres) de précipitations par an, mais les écosystèmes aquatiques sont alimentés par les eaux souterraines.
Le bassin se situe à 750 mètres au-dessus du niveau de la mer, avec les montagnes environnantes s’étendant sur 2500 mètres au-dessus.
Riche en écosystèmes de sources pauvres en azote et en phosphore, le bassin présente un fort taux d’endémisme.
Les micro-organismes présents dans les bassins de Cuatro Ciénegas, comme leurs prédécesseurs à la période précambrienne, la première et la plus longue étape de l’histoire de la Terre, survivent dans un environnement peu nutritif, très différent du reste de la planète.
Bien que dans le passé géologique il y ait probablement eu des connexions avec le Rio Grande, la zone se caractérise aujourd’hui par son isolement.
Des cartes anciennes (1860) montrent que la zone se composait initialement de deux grands lacs, devenus aujourd’hui un fouillis de plans d’eau.
MINCKLEY (1969) a décrit de nombreux aspects de la vallée, tels que la géologie, l’eau et la biologie.
La vallée mesure environ 700 km2, et est située à environ 740 mètres au-dessus du niveau de la mer.
Elle abrite des centaines de sources qui varient en taille de quelques mètres carrés à plusieurs hectares.
Ce sont souvent des sources chaudes avec des températures allant jusqu’à 34°C.
Mais des sources froides jaillissent aussi parfois à quelques mètres des sources chaudes et se jettent ensemble dans les ruisseaux des rivières et des marécages.
La vallée est entourée de crêtes montagneuses.
Dans la vallée, les eaux sont moins séparées qu’il n’y paraît et de nombreux plans d’eau sont reliés entre eux par des rivières souterraines et, en outre, les fortes précipitations peuvent créer des connexions temporaires dans de nombreux endroits.
La déshydratation et les espèces exotiques menacent la vallée.
Beaucoup de ces biotopes isolés du désert, en particulier dans le sud des États-Unis, ont disparu avec le temps.
Mais dans l’eau de cette oasis existe une grande diversité de micro-organismes, semblables à ceux qui existent à la première époque de la planète.
Cuatro Ciénegas est une zone humide située à Coahuila, dans le nord du Mexique, un endroit unique dans le monde.
Le système biologique Quatre Ciénegas est un monde perdu dans le désert de Coahuila ; il a été découvert qu’il permettait la réalisation d’études moléculaires et microbiologiques.
Dans ce milieu, il a été détecté la présence de bactéries vivantes similaires à celles qui existent depuis des millions d’années, au moins 200 bactéries identifiées proviennent de celles qui sont apparus il y a des millions d’années, lorsqu’il n’y avait pas d’oxygène dans l’atmosphère, et selon les scientifiques, ces bactéries pourraient être voir la clé de la « cure » de la planète : .
« Cuatro Ciénegas est le lieu le plus diversifié de la planète ».
Les micro-organismes dans les ressources de Cuatro Ciénega, comme ceux de nos ancêtres dans la période pré-cambrienne, la première et la plus grande étape de l’histoire de la Terre, survivent dans un environnement avec peu de nutriments, très différents du reste de la planète.
L’eau des substances contient un haut contenu en minéraux, elle peut être en contact avec des résidus volcaniques, explique l’expert.
Ses éléments sont comparables à ceux qui existent dans la première phase de la Terre, lorsqu’ils se retrouvent dans un processus d’instabilité.
Aujourd’hui, ce lieu recueillerait plus de 3 000 types de bactéries distinctes.
Il existe également une grande diversité d’insectes, de poissons et d’autres espèces considérées comme uniques dans le monde.
« C’est le site unique sur la terre qui garde les sédiments de ce qui était une planète très primitive ».
Il y a des millions d’années dans la zone où se trouve aujourd’hui l’état de Coahuila, il y a eu une fracture de plaques tectoniques qui est originaire de ce qui est aujourd’hui l’océan Atlantique.
Pendant la période préparatoire, la formation de la planète, comme elle est désormais connue, n’est pas terminée.
La région de Quatre Ciénegas, dans ces conditions s’est alors recouverte d’eau.
Dans un tel cas, les conséquences sont beaucoup plus graves que, par exemple, l’assèchement d’un bras de rivière.
Dans ce dernier cas, il y a tout simplement moins d’habitat pour une espèce, mais dans le premier, un pool génétique complet est éradiqué.
L’eau est souvent extraite pour produire des aliments pour animaux.
L’introduction par libération d’espèces exotiques est également une menace croissante !
Cuatro Ciénegas a été déclarée zone naturelle protégée en 1994.
–
MILIEU NATUREL & BIOTOPE
La vallée de Cuatro ciénegas présente un climat de type désertique, allant de très sec à semi-chaud, avec un très faible pourcentage de précipitations hivernales.
Elle se caractérise par une grande amplitude thermique, avec des températures extrêmes de 0°C à 44°C (MINCKLEY, 1969).
Les rares précipitations varient entre 100 et 440 millimètres par an et se produisent principalement durant l’été (de mai à octobre), sous forme d’averses peu fréquentes.
Les périodes de sécheresse sont relativement courantes.
La température moyenne mensuelle la plus élevée dépasse 30°C, et la plus basse est inférieure à 12 °C (SEMARNAT, 1999).
L’habitat des Herichthys se compose principalement de sources thermales dont l’eau est cristalline et de bassins dont les dimensions n’atteignent généralement pas plus de 20 mètres de diamètre, ces endroits étant entourées par un environnement très aride avec une température de l’eau toujours constante.

Les mojarra (Herichthys ) de Quatre Ciénegas habitent dans une région du nord-est du Mexique qui offre un réseau exceptionnellement flexible pour évaluer les patrons de flux génétiques, la divergence et, par conséquent, les modèles d’espèce.
Cette espèce affectionne les sources thermales aux eaux cristallines (visibilité élevée : 20 mètres ou plus) dont la température se situe entre 30 et 34°C.
Ces sources présentent des températures très stables (variation annuelle inférieure à un degré Celsius), ainsi que des niveaux et une qualité d’eau constants (WINSBOROUGH 1990 ; HENDRICKSON & Al., 2005).
L’espèce est active et abondante dans les mares (de 50 centimètres à 5 mètres de profondeur) aux fonds épais de détritus, où l’eau émerge à travers des fonds rocheux propres (travertin ou fragments de stromatolithes).
Nombre de ces mares contiennent divers stromatolithes qui ressemblent à la roche-mère, et dans d’autres cas, ils ont la forme de certains coraux, comme s’il s’agissait de récifs marins.
En aval, plus loin des sources, dans les ruisseaux et les marais, on peut encore trouver des Herichthys minckleyi, cependant, en hiver, lorsque la température chute drastiquement, la présence de Herichthys minckleyi devient rare, voire inactive ou totalement absente.
Cette espèce de cichlidé n’a pas été observée dans les eaux plus saumâtres et instables de la vallée, comme la Laguna Grande, où les fluctuations de température, de salinité et de niveau d’eau sont généralement importantes.
Différents types de stromatolithes, formés par des communautés variées de diatomées et de cyanobactéries, sont présents dans de nombreux habitats de ces espèces (WINSBOROUGH 1990 ; WINSBOROUGH & Al., 1994 ; ELSER & Al., 2005).
La végétation autour des mares fréquentées par ces espèces est principalement composée de graminées et d’herbes halophytes charnues à petites feuilles, de moins d’un mètre de hauteur.
Parmi les espèces dominantes, on trouve :
- Distichlis spicata ;
- Clappia suaedaefolia ;
- Suaeda mexicana ;
- Sporobolus airoides ;
- Salicornia sp. ;
- Atriplex sp.
-

- Distichlis spicata
-

- Clappia suaedaefolia
-

- Suaeda mexicana
-

- Sporobolus atiroides
-

- Salicornia sp.
-

- Atriplex sp.
Ce cichlidés est originaire du Mexique, Cuatro Cienegas, une petite vallée dans le désert de Chihuahuan.

Par exemple, Herichthys minckleyi se limite à la petite vallée de Cuatro Ciénegas (40 kilomètres x 40 kilomètres) dans le désert chihuahuense mexicain.

Cette espèce fréquente les lagunes, les sources d’eau froide à chaude, les étangs et les ruisseaux aux eaux très claires, parfois légèrement salines.
On rencontre ce cichlidé généralement dans des courants faibles ou nuls, au milieu de débris marneux, de vase, de gravier et de roches, ainsi que d’une végétation immergée comprenant :
- des Chara ;
- des Nymphaea ;
- des Utricularia ;
- des Eleocharis ;
- des Typha.
-

- Scirpus lacustris
-

- Scirpus lacustris
-

- Typha latifolia
-

- Phragmites
-

- Eleocharis palustris
Les espèces aquatiques telles que les nénuphars (Nymphaea) et les Chara se rencontrent dans les mares, elles sont souvent accompagnées de plantes émergentes sur leurs berges, telles que :
- Typha ;
- Scirpus ;
- Phragmites ;
- Eleocharis ;
- ..
Herichthys minckleyi évolue jusqu’à des profondeurs allant jusqu’à 7,0 mètres (MILLER & Al., 2005).

La vallée possède plusieurs petits lacs, dont certains ont des sources thermales.
Au totale il y a 16 sortes de poissons présents dans ces lacs, dont 8 sont endémiques comme l’est Herichthys Minckleyi.
Le fond des lacs, se constitue principalement d’un substrat rocheux.
De temps en temps on trouve des zones ou le fond rocheux est découvert de détritus.
Dans certaines sources on trouve un fond plâtreux.
Dans les rives on trouve des rochers plâtreux, les plantes les plus courantes sont de la famille des Nymphéas.

L’eau à un Ph de 7.6 à 7.8 et la température des sources est très stable.
En effet, des mesures de température effectuées sur dix ans dans La Laguna Del Mojarral montrent des températures qui restaient entre 33,5° et 34°C, ce qui donne au poisson le possibilité de se reproduire durant toute l’année.
Los Mesquites River (qui se trouve uniquement dans la vallée Cuatro Cienegas) et certaines autres lagunes ont des températures moins élevées et des plus grandes différences durant l’année.
Dans cette région, la période de reproduction se situe alors aussi dans les mois plus chauds et secs.
L’habitat de ces poissons est constitué de sources thermales et d’étangs cristallins, à température constante et d’un diamètre d’au plus 20 mètres.
–
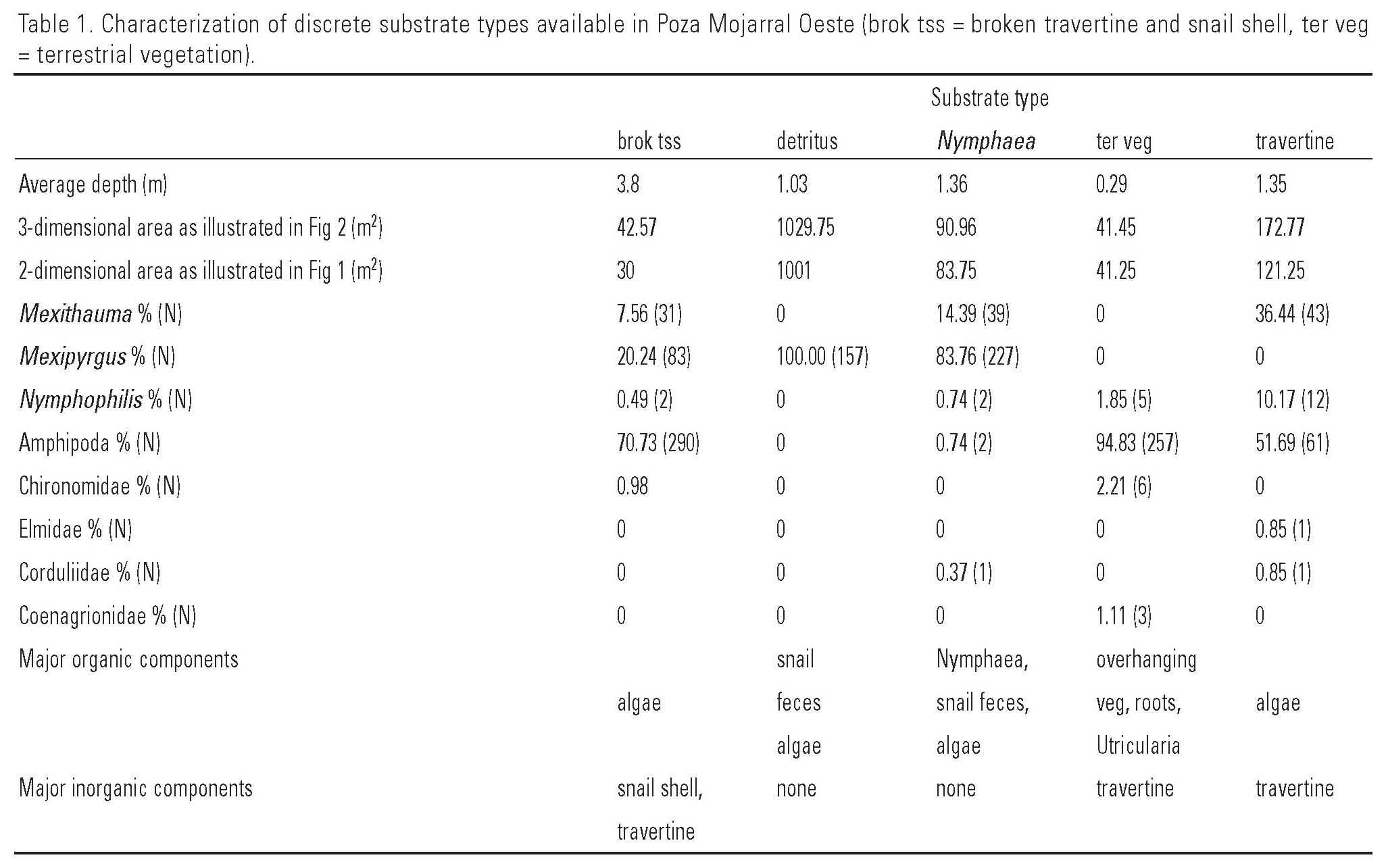
Caractérisation et disponibilité d’un habitat type : La Poza Mojarral Oeste
La Poza (bassin ou piscine naturel) Mojarral Oeste possède 1377,5m2 de surface totale de substrat au fond, des profondeurs allant jusqu’à 4,88 mètres, et 5 types de substrats bien définis :
Le travertin
Le travertin est une formation calcaire biogénique stratifiée avec une faune algale et bactérienne vivante riche dans les couches superficielles (WINSBOROUGH, 1990) qui couvre 12,54 % de la superficie totale des bassins.

Dans certaines parties de la piscine, elle est recouverte d’une fine poussière détritique, mais elle est le plus souvent propre et exposée.
Dans certains endroits, le travertin est associé à des pentes raides atteignant des profondeurs de 0,5 à 4,88 mètres.
Le courant est important dans certaines zones de travertin près des sources de printemps et dans le siphon.
Le travertin dans ce bassin présente une surface très irrégulière, avec de nombreuses crevasses et conduits de tailles variées largement utilisés comme couverture par Herichthys minckleyi et d’autres poissons.
–
La zone des détritus
Cette zone est composée presque entièrement de crottes d’escargot et de l’escargot Mexipyrgus churinceanus (TAYLOR, 1966).
Cette substance floculante claire, facilement perturbée pour troubler l’eau, couvre 74,75 % du substrat total de la « piscine ».

Les densités d’escargots sont plus élevées dans ce substrat que dans tout autre et bien qu’ils vivent dans ce matériau floculant, ils ne sont généralement pas visibles à sa surface.
La zone des Nymphéas
Le Nymphaea est un nénuphar « Nymphaea ampla » (Salisbury) de Candolle qui présente deux formes de croissance :
- L’une avec des feuilles flottantes ;
- L’autre avec des feuilles submergées et ébouriffées près du fond.

La forme submergée prédomine dans Poza Mojarral Oeste, couvrant 6,6 % de la superficie totale du bassin.
–
La végétation terrestre
La végétation terrestre est composée de racines, feuilles et branches de végétation terrestre qui recouvrent les berges et déploient une grande partie du bord du bassin.
Des espèces aquatiques telles que Scirpus sp. y sont présentes et Utricularia sp. est souvent emmêlée dans les racines submergées.
Cette zone de surplomb offre un bon abri pour les poissons et représente 3,01 % de la surface totale du bassin.
–
Une zone mixte
Cette zone est composée de travertin[1] brisé et de coquilles d’escargots.
Elle se trouve dans les endroits les plus profonds, principalement dans deux zones correspondant à des vitesses de courant élevées qui représentent ensemble 3,09 % de la surface totale du bassin.

Ce substrat se compose de fragments de travertin qui se sont détachés des formations de travertin plus grandes en surplomb, ainsi que d’un ensemble de coquilles d’escargots morts, principalement du Mexipyrgus, mais aussi un peu de Mexithauma.
Les types de substrats diffèrent bien plus que par leurs caractéristiques physiques simples (Tableau 1).
{kind=link}

Tous les substrats, à l’exception de Nymphaea et de travertin, diffèrent significativement dans la profondeur moyenne à laquelle ils se trouvent (ANOVA, p<0,0001), allant de quelques centimètres pour la végétation terrestre à jusqu’à 4,88 mètres de profondeur pour le travertin cassé et la coquille d’escargot.
Le seul invertébré recensé dans les débris est Mexipyrgus churinceanus.

Mexipyrgus churinceanus.
Dans le travertin brisé et la coquille d’escargot :
- Mexipyrgus churinceanus représente environ 20 % de la faune d’invertébrés ;
- de Hyalella sp. représente environ 70 %.

Hyalella sp.
Au sein de Nymphaea :
- Mexipyrgus churinceanus représente environ 84 % de la faune d’invertébrés ;
- Mexithauma quadripaludium (TAYLOR, 1966) représente 14 %.

Mexithauma quadripaludium.
Dans la végétation terrestre de Hyalella sp. représente 95 % du total des invertébrés.
Dans le travertin :
- Hyalella sp. [2]représente environ 52 % des invertébrés ;
- Mexithauma quadripaludium [3]représente environ 36 % ;
- Nymphophilus minckleyi représente 10 %.

Nymphophilus minckleyi.
D’autres insectes aquatiques se trouvent également dans certains substrats, notamment des membres des familles :
- Chironomidae (Diptera) ;
- Elmidae (Coleoptera) ;
- Corduliidae ;
- Coenagrionidae (Odonata), bien que dans de faibles densités.
-

- Chironomidae Diptera.
-

- Chironomidae Diptera. (larve)
-

- Elmidae Coleoptera ( larve)
-

- Elmidae Coleoptera
-

- Corduliidae somatochlora
-

- Coenagrionidae
Plusieurs autres poissons se trouvent également dans ces environnements :
- Cyprinodon bifasciatus – MILLER, 1968 – très courants ;
- Gambusia marshi – MINCKLEY & CRADDOCK, 1962 – très courants ;
- Cyprinella xanthicara – MINCKLEY & LYTLE, 1969 – présents, mais peu communs ;
- Astyanax mexicanus – DE FILIPPI, 1853 – présents, mais peu communs ;
- Ictalurus lupus – GIRARD, 1858 – présents, mais peu communs.
-

- Astyanax mexicanus
-

- Ictalurus lupus
-

- Cyprinella xanthicara
-

- Gambusia marshi
-

- Cyprinodon bifasciatus
–
État actuel de l’habitat au regard des besoins de l’espèce
La plupart des mares où les premières collectes ont été effectuées dans les années 1960 et 1970 sont actuellement en bon état.
Cependant, la vallée a subi des modifications importantes qui affectent la répartition des habitats aquatiques (MINCKLEY 1969, 1992 ; HENDRICKSON & Al. 2005).
La rivière Garabatal, autrefois très importante et prenant sa source dans la mare de La Becerra, est aujourd’hui à sec ou presque, car son débit a été détourné dans les années 1960 à des fins d’irrigation (HENDRICKSON & Al., 2005).
Bien que l’espèce soit encore présente dans les habitats résiduels de ce système fluvial, la population la plus importante, qui occupait jadis de vastes marais, rivières et mares, est actuellement très fragmentée, clairsemée et risque fortement de disparaître dans un avenir proche (HENDRICKSON & Al., 2005).
À la Poza de la Becerra, toutes les formes de Herichthys minckleyi sont encore présentes, cependant, les populations subissent une forte pression car leur habitat est affecté par le tourisme et les fluctuations du niveau d’eau liées à la gestion du système d’irrigation.
Les stromatolithes[4], autrefois communs dans le bassin, ont disparu à cause de l’activité humaine dans ce complexe touristique.

Hormis la limnée tuberculée (Melanoides tuberculata), aucune espèce invasive n’est présente dans le système.
Certaines espèces autrefois recensées, comme l’Achigan à grande bouche (Micropterus salmoides ssp.), ont déjà disparu.

Micropterus salmoides ssp. : Cette espèce est considérée comme l’une des espèces exotiques les plus nuisibles au monde car elle est carnivore, extrêmement vorace et s’attaque à pratiquement tous les invertébrés et vertébrés qu’elle peut ingérer. De plus, sa reproduction est très prolifique (les mâles prennent soin des œufs, pondus par une ou plusieurs femelles, parfois plus de 10 000 œufs par femelle). Ceci a entraîné l’extinction d’autres populations animales, par prédation directe ou par compétition trophique, et a eu un effet dévastateur, notamment sur les populations de poissons indigènes. Des études indiquent qu’elle possède une grande plasticité trophique, ce qui signifie qu’elle peut modifier son régime alimentaire pour survivre. Son impact écologique est donc évident car elle élimine de nombreux éléments des réseaux trophiques des écosystèmes aquatiques, provoquant la disparition d’espèces indigènes moins prisées pour la pêche mais essentielles à la biodiversité locale. Pour ces raisons, cette espèce figure sur la liste des 100 espèces exotiques envahissantes les plus nuisibles au monde, selon l’UICN.
La disparition du fleuve Garabatal dans la vallée de Cuatro Ciénegas est l’un des impacts fauniques les plus importants enregistrés à ce jour.
L’espèce est également menacée par des espèces exotiques, comme Hemichromis guttatus, un cichlidé africain d’origine qui s’est largement diversifié pour occuper l’Amérique centrale et du sud. (CONTRERAS-BALDERAS & LUDLOW 2003 ; HENDRICKSON & Al. 2005).

Bien évidemment, il s’est aussi établi dans presque tous les habitats d’Herichthys minckleyi.
Des études d’isotopes stables indiquent que le régime alimentaire de Herichthys guttatus présente un chevauchement significatif avec celui des juvéniles de Herichthys minckleyi et des adultes de Cyprinodon bifasciatus, une autre espèce endémique des sources thermales (HENDRICKSON & Al., 2005).

Cyprinodon bifasciatus.
Le tilapia (Oreochromis aureus) est abondant dans la partie sud-est de la vallée et a été observé dans le cours moyen et inférieur du bassin versant de la rivière Mezquites (HENDRICKSON & Al., 2005).

Oreochromis aureus.
Parmi les autres menaces potentielles figurent sans aucun doute l’escargot invasif Melanoides tuberculata, établi dans le canal de Becerra et le bassin de Churince, ainsi que l’écrevisse exotique Procambarus clarkii (présente dans les rivières MEZQUITES & GARABATAL) (HENDRICKSON & Al., 2005).

Melanoides tuberculata.

Procambarus clarkii.
Il ne faut pas oublier les plantes envahissantes Arundo donax et Tamarix ramosissima (HENDRICKSON & Al., 2005 ; HENDRICKSON & McGAUGH, 2005).

Arundo donax.

Tamarix ramosissima.
–
Utilisation de l’habitat
L’espèce privilégie les sources chaudes et les zones situées à proximité de l’aval, cherchant apparemment refuge dans des milieux plus chauds durant l’hiver.
Au sein des bassins, des études récentes ont mis en évidence des préférences de substrat distinctes selon les différents morphotypes, ainsi que des variations saisonnières (SWANSON 2001 ; SWANSON & AL. 2003 ; COHEN & AL. 2005).
La température dans la zone de nidification se situe généralement entre 29 et 30°C.
Suite à une panne du thermostat de chauffage, d’un aquarium quatre individus qui y étaient présents, ont été retrouvés vivants et apparemment à leur aise à 40°C.

Cette expérience malheureuse démontre une certaine capacité de ces poissons à résister à des températures extrêmes…au moins pour un temps réduit !
Dans des étangs expérimentaux, où l’eau est à une température de 30 (± 2°C d’écart) pendant au moins six mois de l’année, de nombreux individus ont survécu plus de trois jours à des températures inférieures à 12°C, dont 20 heures consécutives à des températures comprises entre 10,9 et 11,5°C.
En revanche, à partir d’expériences faites en aquarium, il a été observé que l’activité de l’espèce est faible à des températures inférieures à 20°C.
Cependant, des populations maintenues dans des bassins à ciel ouvert ont survécu à des températures aussi basses que 10°C.
Les variations d’abondance enregistrées dans certains bassins durant l’hiver indiquent des déplacements probables de certains individus (ou une absence d’activité dans certaines zones), possiblement pour se réfugier près des sources thermales.
Toutefois, des études de marquage-recapture n’ont enregistré aucun mouvement entre deux bassins voisins (Mojarral Oeste et Mojarral Este), qui présentent une différence de température de 1 à 3°C, malgré le marquage et le lâcher de milliers de spécimens (obs. pers.).
Ce résultat suggère (à titre préliminaire) que la population du bassin thermal est relativement fermée.
Le programme de marquage-recapture mené pendant six ans à Mojarral Oeste (Poza Azul), impliquant de multiples captures (jusqu’à 11 fois) de nombreux individus, indique qu’au moins certains (et probablement la plupart) des individus ne quittent jamais, ou rarement, ce bassin thermal.
De plus, la proportion d’individus marqués varie de 40 à 70 %, avec quelques individus adultes non marqués trouvés dans presque tous les échantillons, ce qui indique probablement une faible immigration.
TAXONOMIE
HISTOIRE NATURELLE DE L’ESPECE
Contexte sur le statut de l’espèce et des principales populations
On ne dispose pas de données précises sur la taille des populations, mais les observations des collecteurs et les captures indiquent une abondance sur de nombreux sites de collecte.
–
Cycle de vie
L’espèce est connue pour son polymorphisme (SAGE & SELANDER 1975 ; KORNFIELD 1981 ; KORNFIELD & AL., 1982 ; KORNFIELD & TAYLOR, 1983).
Au sein de cette même espèce, on observe trois types morphologiques (« morphes ») si distincts que leurs premiers auteurs les ont considérés comme des espèces séparées (TAYLOR & MINCKLEY 1966 ; LABOUNTY 1974 ; MINCKLEY 1969 ; KORNFIELD & KOEHN 1975).
Cependant, des études génétiques ultérieures n’ont pas mis en évidence de différences génétiques entre les morphes et ont démontré l’absence d’isolement sexuel entre eux.
Le polymorphisme principal concerne les dents pharyngiennes, c’est-à-dire les dents internes de la gorge, et n’est donc pas très visible de l’extérieur.
Cependant, à l’examen de ces dents, certains individus présentent des dents très grandes et robustes (molaires), tandis que d’autres ont des dents très fines, pointues et délicates (papillaires).
Les dents molaires sont ancrées dans des os épais et résistants, associés à des muscles puissants et développés. Les individus aux dents papillaires ont des os pharyngiens beaucoup plus fins et des muscles associés moins développés (LIEM & KAUFMAN, 1984).
Les individus aux dents molaires consomment des escargots, contrairement à ceux aux dents papillaires. Les escargots endémiques de Cuatro Ciénegas, diversifiés et abondants, appartenant à la famille des Hydrobiidae (HERSHLER 1984, 1985 ; HERSHLER & MINCKLEY, 1986 ; HERSHLER & HAYEK, 1988), possèdent les coquilles les plus résistantes de toute la planète (Hulsey & Al., 2005a).
Il est probable que cette dureté ait été acquise par coévolution avec leur prédateur, Herichthys minckleyi, autrement dit, l’évolution de la forme molariforme est liée à la prédation sur les escargots à coquille aussi dure.
Les individus à dents papillaires ne peuvent briser la coquille de ces escargots et, par conséquent, ne peuvent ni les exploiter ni les consommer.
En lien avec ces différences alimentaires, les individus à dents papillaires possèdent des intestins deux à quatre fois plus longs que ceux de même taille à dents molariformes (LABOUNTY 1974 ; KORNFIELD & KOEHN 1975).
Les premiers chercheurs ont rapporté une faible proportion d’individus présentant une morphologie dentaire pharyngienne intermédiaire, mais la quantification et la description généralement insuffisantes de cette morphologie compliquent la comparaison avec les données historiques.
STEPHENS (2002) a examiné de nombreux spécimens étudiés par ces premiers chercheurs et a constaté, dans les collections récentes, une proportion plus élevée d’individus à dents intermédiaires.
Il a suggéré que cette proportion avait augmenté ces dernières années, peut-être en raison de changements environnementaux, et a considéré ces individus comme un autre morphotype distinct du système polymorphe.
Des études génétiques à plus haute résolution sont en cours afin de répondre à ces questions et à d’autres concernant les détails de l’évolution et du maintien du polymorphisme (GARCIA DE LEON, données non publiées ; HENDRICKSON, données non publiées ; HULSEY, données non publiées).
Bien que les études soient toujours en cours, il est probable qu’un autre morphotype existe, se distinguant des autres par la forme de son corps et de sa tête.
Alors que les individus des morphotypes principaux ont un corps et une tête de forme « normale », ce morphotype, appelé « piscivore », présente une tête et un corps beaucoup plus allongés.
Son régime alimentaire comprend des poissons, notamment Cyprinodon bifasciatus.

Les piscivores peuvent présenter des dents pharyngiennes molariformes, mais cette combinaison est rare, et la plupart des individus à morphologie corporelle et céphalique de type piscivore possèdent des dents papilliformes (D. HENDRICKSON, observation personnelle).
Les deux principaux morphotypes « normaux » sont largement sympatriques, tandis que les piscivores sont principalement confinés aux zones proches des sources thermales.
Ces derniers adoptent une coloration cryptique à bandes verticales qui leur permet d’utiliser une technique de chasse à l’affût contre Cyprinodon.
Des études en laboratoire indiquent que la morphologie des dents pharyngiennes est déterminée en partie par des facteurs génétiques, mais aussi par des facteurs environnementaux (TRAPANI, 2001, 2003, 2004 ; STEPHENS, 2002).
Les proportions des différents morphotypes varient selon les bassins de la vallée et au fil du temps (STEPHENS, 2002).
–
Pertinence de l’espèce
Pertinence écologique
L’abondance et la diversité de son régime alimentaire témoignent du rôle essentiel de cette espèce dans le fonctionnement des écosystèmes thermaux de la vallée de Cuatro Ciénegas.
L’impact des espèces d’escargots introduites sur les stromatolithes est bien documenté (ELSER & Al., 2002, 2005).
L’absence d’Herichthys minckleyi favoriserait probablement l’augmentation des populations d’escargots sauvages, ce qui aurait un impact négatif sur l’ensemble de la chaîne alimentaire des seuls écosystèmes aquatiques de la vallée.
–
Pertinence taxonomique
Les premiers ichtyologues ayant collecté l’espèce ont considéré ses différents morphotypes comme des espèces distinctes.
Cependant, des études génétiques ont démontré l’absence de différences entre eux, et H. minckleyi a donc été décrite comme une espèce polymorphe.
Des données génétiques préliminaires plus détaillées que celles des études initiales, mais non encore publiées, confortent l’hypothèse présentée dans la description, selon laquelle l’espèce représente un complexe d’espèces naissant dans le processus de spéciation actuel.
Par conséquent, cette espèce est considérée comme très importante pour les études sur le processus de spéciation dans la région.
–
Importance économique
Elle n’a pas de valeur économique directe, mais présente une grande valeur économique indirecte pour la recherche fondamentale et l’écotourisme.
L’espèce est très visible et bien connue de tous les touristes et à ce titre, elle est utilisée à des fins commerciales de toutes sortes puisqu’elle apparaît fréquemment dans l’art populaire et sur les articles vendus aux touristes locaux (T-shirts, sculptures, etc.).
–
Importance culturelle
Sa grande visibilité dans les eaux cristallines des bassins et ses couleurs ont fait de cette espèce l’un des symboles les plus connus (avec la tortue « Terrapene coahuila ») de la faune endémique de la vallée.

Terrapene coahuila.
Tous les habitants et les touristes de la vallée la connaissent bien.
Avant l’interdiction de la pêche imposée après la déclaration de l’aire protégée, elle était fréquemment capturée.
On entend souvent dire que « sa chair est la meilleure du monde », et la « mojarra frite de Cuatro Ciénegas » était considérée comme un mets de choix.

–
Herichthys Minckleyi (KORNFIELD & TAYLOR, 1983)
- Basionyme : Cichlasoma minckleyi (KORNFIELD & TAYLOR, 1983) dans [KORNFIELD & TAYLOR JN (1983)]
Holotype de Cichlasoma Minckleyi
- UMMZ : 209434

Paratype de Cichlasoma Minckleyi :
- USNM : 229347 ;
- USNM : 231944-47

- Paratype UMMZ 18132 : Mâle Herichthys minckleyi, 93.5 millimètres. Morphe papiliforme issu de la Pozos de la Becerra, Cuatro Ciénegas, 1961.

- Paratype UMMZ 179832 : Mâle 93,5 mm LP. Forme avec dents pharyngiennes papillaires, Poza de la Becerra, Cuatro Ciénegas de Carranza, Coahuila, Mexique, 1961 (KORNFIELD & TAYLOR, 1961).

Paratype UMMZ 209434 : Mâle Herichthys minckleyi, Holotype, Morphe papiliforme. Pos de la Becerra, Cuatro Ciénagas, 1961.
–
Localité type
- Posos de la Becerra, à 15,7 kilomètres par la route au sud-sud-ouest de Cuatro Ciénegas de Carranza, Coahuila, Mexique.
–
Synonymes
Cichlasoma Minckleyi (KORNFIELD & TAYLOR, 1983).
–
RAPPEL SUR LES …
Informations sur le genre Herichthys
Les cichlidés du genre Herichthys sont très répandus dans le nord-est du Mexique et le sud du Texas (bassin hydrographique du Rio Grande).
Tous les Herichthys sont des cichlidés robustes avec un corps comprimé latéralement et un dimorphisme sexuel modéré.
La plus petite espèce des Herichthys est Herichthys deppii avec une longueur standard maximale d’environ 12 centimètres tandis que la plus grande espèce est Herichthys steindachneri, qui atteindrait des longueurs standard pouvant atteindre 40 centimètres.
Le reste se situe confortablement entre ces deux extrêmes, et dans des conditions d’aquarium, au moins Herichthys cyanoguttatus et Herichthys carpintis atteignent tous deux entre 20 et 30 centimètres de longueur, les mâles étant généralement plus gros que les femelles.
Les différentes espèces de Herichthys affichent tous des motifs de couleurs distinctifs. Cependant, un thème commun est la façon dont ils ajustent leurs couleurs avec l’humeur, en particulier lors de la reproduction.
De vastes motifs sombres sont souvent observés sur les flancs des spécimens reproducteurs, le mâle et la femelle affichant des couleurs similaires.
Il y a cependant un certain dimorphisme sexuel.
En plus d’être légèrement plus gros que la femelle, les Herichthys mâles ont souvent des nageoires dorsale et anale visiblement plus longues et plus pointues, et les mâles plus âgés peuvent développer une bosse nucale en vieillissant.
Jusqu’à 2013, la systématique au niveau du genre « Herichthys » était stable et comprenait neuf espèces.
Au cours des dernières années, le genre Herichthys a connu plusieurs bouleversements taxonomiques importants qui comprenait les descriptions formelles de plusieurs espèces et la ségrégation de certaines espèces qui ont été placées dans de nouveaux genres.
Dans un premier temps, DE LA MAZA-BENIGNOS et LOZANO-VILANO (2013) et DE LA MAZA-BENIGNOS & Al. (2015) ont décrit trois espèces auparavant de manière informelle connu sous le nom de :
- « Labridens blancs » (Herichthys pame) ;
- « Labridens verts » (Herichthys pratinus – DE LA MAZA-BENIGNOS & LOZANO-VILANO, 2013) ;
- « Turquoise Herichthys » (Herichthys tepehua).
…plus une espèce, probablement nouvelle : Herichthys molango.

Herichthys molango.
DE LA MAZA-BENIGNOS & LOZANO-VILANO, 2013 ont ensuite élevé « Herichthys teporatus » (FOWLER 1903) au rang d’espèce valide.
Mais, il y a un problème avec toutes ces descriptions d’espèces car elles manquent de caractères les diagnostiquant et en plus n’étaient pas prises en charge par le présenté (ADNmt) phylogénies.
Deuxièmement, DE LA MAZA-BENIGNOS & Al. (2015) ont séparé les espèces anciennement incluses dans le groupe d’espèces « Herichthys labridens » en créant un nouveau genre nommé « Nosferatu », encore une fois malgré le manque de monophylie dans leur phylogénie d’ADNmt et avec une question morphologique possible qui contredit en outre la seule phylogénie morphologique du genre disponible (RICAN & al., 2008).
Le genre Herichthys a, ainsi, subi récemment plusieurs changements taxonomiques généraux qui ne sont pas pris en charge par les données et sont, dans de nombreux cas, réfutés par des données supplémentaires (MEJIA, PEREZ-MIRANDA, LEON-ROMERO, SOTO-GALERA, & DE LUNA, 2015 ; RICAN & Al., 2016).

Le genre Herichthys compte actuellement 11 espèces connues et décrites (http: //aquatab.net / …) :
- Herichthys bartoni ;
- Herichthys carpintis ;
- Herichthys cyanoguttatus ;
- Herichthys dépii ;
- Herichthys labridens ;
- Herichthys minckleyi ;
- Herichthys pantostictus ;
- Herichthys steindachneri ;
- Herichthys tamasopoensis ;
* les espèces désignées sont inscrites sur la liste rouge de l’UICN des espèces menacées d’extinction depuis 1996.
KULLANDER dans REIS & Al. (2003) mentionne 10 espèces du genre Herichthys:
- Herichthys deppii – HECKEL, 1840 ;
- Herichthys geddesi – REGAN, 1905 « incertae cedis » ;
- Herichthys carpintis – JORDAN & SNYDER, 1899 ;
- Herichthys pantostictus – TAYLOR & MILLER, 1983 ;
- Herichthys tamasopoensis – ARTIGAS-AZAS, 1993 ;
- Herichthys steindachneri – JORDAN & SNYDER, 1899 ;
- Herichthys bartoni – BEAN, 1892 ;
- Herichthys labridens – PELLEGRIN, 1903 ;
- Herichthys cyanoguttatus – BAIRD & GIRARD, 1854.
- Herichthys minckleyi – KORNFIELD ;
Considéré comme synonyme de « Cichlasoma » pendant plusieurs décennies, « Herichthys » compte actuellement une dizaine d’espèces, toutes originaires d’Amérique centrale.
Selon FishBase (25 janv. 2017) :
- Herichthys carpintis – JORDAN & SNYDER, 1899 ;
- Herichthys cyanoguttatus – BAIRD & GIRARD, 1854 ;
- Herichthys deppii – HECKEL, 1840) ;
- Herichthys minckleyi – KORNFIELD &TAYLOR, 1983 ;
- Herichthys tamasopoensis – ARTIGAS AZAS, 1993 ;
- Herichthys tepehua – DE LA MAZA-BENIGNOS, ORNELAS-GARCIA, LOZANO-VILANO, GARCIA-RAMIREZ & DOADRIO,
Selon World Register of Marine Species (25 janv. 2017) :
- Herichthys pantostictus – TAYLOR & MILLER, 1983.
Selon ITIS (25 janv. 2017) (non mis à jour depuis 2004) :
- Herichthys bartoni – BEAN, 1892 ;
- Herichthys carpintis – JORDAN & SNYDER, 1899 ;
- Herichthys cyanoguttatum – BAIRD & GIRARD, 1854 ;
- Herichthys deppii – HECKEL, 1840 ;
- Herichthys labridens – PELLEGRIN, 1903 ;
- Herichthys minckleyi – KORNFIELD & TAYLOR, 1983 ;
- Herichthys pantostictus – TAYLOR & MILLER, 1983 ;
- Herichthys pearsei – HUBBS, 1936 ;
- Herichthys steindachneri – JORDAN & Snyder, 1899 ;
- Herichthys tamasopoensis – ARTIGAS AZAS, 1993.
Plusieurs autres espèces ont été proposées récemment, ainsi :
- Herichthys pratinus – DE LA MAZA BENIGNOS & LOZANO-VILANO, 2013 ;
- Herichthys pame – DE LA MAZA BENIGNOS & LOZANO-VILANO, 2013 ;
- Herichthys molango – DE LA MAZA BENIGNOS & LOZANO-VILANO, 2013.

(a) Herichthys bartoni
(b) Herichthys labridens
(c) Herichthys pame
(d) Herichthys pantostictus (lacustre population)
(e) Herichthys carpintis
(f) Herichthys tepehua
(g) Herichthys tamasopoensis
(h) Herichthys deppii
(i) Herichthys cyanoguttatus,
(j) Herichthys minckleyi (femelle ; le mâle est tout noir)
Herichthys est divisé en deux groupes monophylétiques :
- Premier groupe, dit « labridens » comprenant :
- Herichthys bartoni ;
- Herichthys labridens ;
- Herichthys steindachneri ;
- Herichthys pame ;
- Herichthys pantostictus (TAYLOR & MILLER, 1983).
- Deuxième groupe appelé « cyanoguttatus » comprenant :
- Herichthys cyanoguttatus – BAIRD & GIRARD,1854 ;
- Herichthys carpintis – JORDAN & SNYDER, 1899 ;
- Herichthys minckleyi ;
- Herichthys tamasopoensis – ARTIGAS AZAS, 1993 ;
- Herichthys tepehua – DE LA MAZA-BENIGNOS, ORNELAS-GARCIA, LOZANO-VILANO, GARCIA-RAMIREZ & DOADRIO, 2015 ;
- Herichthys deppii – HECKEL, 1840.

 Herichthys tamasopoensis se distingue des autres Herichthys qui lui sont le plus semblables, et est plus étroitement apparenté et parapatrique avec Herichthys carpintis par :
Herichthys tamasopoensis se distingue des autres Herichthys qui lui sont le plus semblables, et est plus étroitement apparenté et parapatrique avec Herichthys carpintis par :
- Comparaison avec des zones noircies sur la tête limitée à la partie ventrale de bouche à l’exclusion des lèvres ;
- La présence d’un préopercule tout blanc dans la face latérale interrompant la zone antérieure et postérieure noircie, qui comprend la partie ventrale du sous-opercule et la partie ventrale de l’ensemble du corps combiné avec des barres corporelles postérieures noires ;.
- La coloration nuptiale sur la tête est également un indice Vs Herichthys deppii qui possède des lèvres blanches.
Cette situation est intermédiaire entre Herichthys carpintis, Herichthys cyanoguttatus et Herichthys minckleyi, qui tous trois sont dépourvus de cette partie ventrale noircie de la bouche.
Herichthys tamasopoensis se distingue du reste des espèces du groupe d’espèces Herichthys cyanoguttatus par le fait qu’il possède 13 rayons contre 14 rayons au niveau de la nageoire pectorale.
La coloration de reproduction chez Herichthys est composée d’un noircissement de la partie ventrale de la tête, partie ventrale du corps antérieur et toute la partie postérieure du corps, tandis que le reste la tête et le corps deviennent blancs comme neige (ou jaunes dans un seul cas : Herichthys labridens).
Les modèles de coloration de reproduction spécifiques à l’espèce sont observés dans en détails et en fonction de :
- L’étendue du noircissement sur la tête
→ Noircissement complètement absent chez Herichthys minckleyi et Herichthys cyanoguttatus ;
→ Noircissement maximum dans le groupe Herichthys labridens et Herichthys bartoni.
- Selon les motifs de noircissement sur le corps (sous forme de barres verticales ou de zones noires) :
→ La première typique dans le groupe Herichthys cyanoguttatus ;
→ La seconde dans le groupe Herichthys labridens plus Herichthys bartoni et dans une large mesure également dans Herichthys carpintis ) et les nageoires ventrales.
Herichthys tamasopoensis diffère de Herichthys carpintis par :
- Son pédoncule caudal plus long (moyenne 17, SD 1 Vs moyenne 15, SD 1) ;
- Sa mâchoire inférieure plus courte (moyenne 29, SD 2 Vs moyenne 31, SD 2).
Herichthys tamasopoensis se distingue de façon relative d’Herichthys carpintis, hors coloration de reproduction, par les taches opalescentes qu’il a placés sur le corps et sa tête beaucoup plus petite et plus blanchâtre.
Herichthys tamasopoensis diffère de toutes les autres espèces d’Herichthys par les caractères autapomorphies suivantes :
- La présence de dents de devant rapprochées, tronquées, aplaties, unicuspides à faiblement bicuspides dans les deux mâchoires ;
- La présence de dents latérales bicuspides ;
- Des arcades supérieures et inférieures très bombées ;
- Une plaque pharyngée inférieure robuste, large et rugueuse,
- Deux rangées de molaires papilliformes, de haute densité et non pigmentées sur la ligne médiane de la surface occlusale ;
- Le profil dorsal convexe nettement incurvé entre la région nucale et le premier radius dorsal.
–
Le groupe d’espèces « Herichthys labridens »
Toutes ces espèces se trouvent dans le bassin fluvial de Pánuco.
Herichthys bartoni se distingue de tous les autres espèces Herichthys par une coloration de reproduction composée de toute la partie suborbitale de la tête et les deux tiers aux trois quarts des flancs du corps uniformément noir, tandis que le reste de la partie dorsale du corps est blanc comme neige.
Herichthys bartoni se distingue des autres espèces du groupe d’espèces Herichthys labridens par :
- La présence de marques opalescentes (uniquement présentes chez les espèces du groupe Herichthys cyanoguttatus) sur :
- La partie postérieure du corps ;
- Le pédoncule caudal ;
- les nageoires impaires (en particulier chez les individus reproducteurs).
- L’absence du marquage rouge-violet sur les parties nues (sans écailles) qui établit normalement une zone colorée au niveau de l’aisselle et de la nageoire pectorale. Ce caractère est présent pour toutes les autres espèces du groupe Herichthys labridens.
Herichthys bartoni a un ensemble de taches de couleur rouille sur la zone postérieure de la tête et ante-partie supérieure du corps, y compris l’aisselle de la nageoire pectorale, mais la tâche à l’aisselle est sur une zone écailleuse et n’est donc pas homologue à la zone nue de couleur rouge présente dans le groupe d’espèces Herichthys labridens .
Les mêmes points de couleur rouille sont trouvés le long de la limite entre la tête et le corps et non associés à une zone nue à la base des nageoires pectorales, à la différence de Herichthys bartoni, Herichthys deppii et de certains Herichthys tepehua.
Herichthys bartoni se distingue nettement de Herichthys labridens par :
- Le croisement des couleurs noir & blanc vs noir & jaune coloration (unique pour Herichthys labridens) où la partie noire chez Herichthys bartoni comprend toute la tête en partie ventrale et tout le corps dans sa partie ventrale ;
- L’absence au niveau de la mâchoire pharyngée inférieure, de molaires robustes ;
- Une tête plus grosse avec une bouche plus large ;
- Une mâchoire inférieure prognathe[5] (vs. isognathes ou hypognathe avec la lèvre supérieure dépassant la lèvre inférieure) ;
- La présence de petits points bruns à noirs et largement séparés sur la tête, joue et séries operculaires au lieu de points beaucoup plus rapprochés et souvent des lignes noires vermiculées (ou pas de points du tout) sur un fond bleu-vert fond avec des lèvres bleues, et en manquant toujours de coloration bleue dans nageoires impaires (les nageoires sont orange à rouille).
Herichthys bartoni se distingue également de toutes les autres espèces d’Herichthys en étant la seule espèce à posséder seulement quatre épines de la nageoire anale.
La barre d’Herichthys bartoni est sympatrique et syntopique avec Herichthys labridens.
Cette barre est endémique aux espèces issues des lagunes de la zone Laguna de la Media Luna (San Luis Potosı) qui sont situées à des altitudes comprises entre 1 000 et 1 100 mètres d’altitude et aussi (bien que beaucoup plus rare) dans certaines parties du Haut Rio Verde.
Herichthys labridens diffère totalement de toutes les espèces de Herichthys pour sa couleur de reproduction nuptiale qui est blanche et noire en opposition à la combinaison de colorations jaune et noire de ces autres espèces.
Par rapport aux autres Herichthys, Herichthys tamasopoensis a :
- Un corps moins profond ;
- Un pédoncule caudal plus long ;
- Un museau plus court.
Il y a également un manque de concavité au-dessus de l’œil, présent chez Herichthys carpintis adulte de la localité type.
Herichthys tamasopoensis est également plus petit que Herichthys carpintis, le premier atteignant un maximum de 18 centimètres, tandis que le second mesure 22 centimètres.
Le poisson a été décrit dans le genre Herichthys du groupe Herichthys cyanoguttatus (cichlidés Texas), avec lequel il partage des caractéristiques diagnostiques telles que les dents antérieures de la série externe dans chaque mâchoire qui sont bicuspides et compressées, devenant tronquées chez les adultes.
Les autres caractéristiques comprennent des taches noires sur la moitié antérieure des flancs qui se transforment en un motif de couleur de reproduction, la moitié antérieure étant blanche contrairement aux barres noires alternées sur la moitié postérieure des flancs, avec une variation de Herichthys minckleyi.
Les espèces Herichthys de ce groupe ont également des taches nacrées sur les côtés et les nageoires des poissons de couleur normale.
Ceux-ci s’intensifient chez les individus dominants et varient d’une espèce à l’autre et de la couleur jaune au bleu avec le vert intermédiaire.
Les dents les plus antérieures de la mâchoire supérieure sont spatulées, en forme de ciseau ou prémolaires.
Herichthys tamasopoensis a un corps moins profond, un pédoncule caudal plus long et un museau plus court que le reste des Herichthys.
Les couples changent de coloration grise de base avec six à sept taches noires dans la moitié arrière de leurs flancs dès que les territoires sont formés.
La couleur de base est d’un blanc crème brillant et le noir est présent dans six ou sept barres verticales sur la moitié arrière de leurs flancs, ainsi que dans une bande dans la zone ventrale de la lèvre inférieure à la base de la nageoire anale.
Les nageoires pelviennes deviennent noir velouté.
Une teinte saumon apparaît sur la nageoire dorsale frontale et les branchies.
–
CLES DES ESPECES

CLE DU GENRE HERICHTYINE
AVERTISSEMENT : La plupart des caractères ne s’appliquent qu’aux spécimens adultes.
1 . La nageoire anale possède seulement quatre épines ;
Coloration nuptiale et de reproduction des femelles : Elle est noire continue sur la partie inférieure de la tête et du corps et blanche continue sur la partie supérieure de la tête et du corps ;
Petits points bruns à noirs largement séparés sur la tête, les joues et les séries operculaires ;
Tête et bouche grandes, mâchoire inférieure dépassant devant la mâchoire supérieure. ⇒ Herichthys bartoni

Couple Herichthys bartoni.
(Espèce endémique aux lagons de la zone de la Laguna de la Media Luna (San Luis Potosl) à des altitudes comprises entre 1 000 et 1 100 mètres d’altitude) .
1′. Nageoire anale avec six à sept épines ; points orange/brun rouille assez gros et bien visibles sur le visage, les joues et les côtés antérieurs du corps ;
Les marques bleues trouvées chez d’autres espèces du groupe d’espèces Herichthys cyanoguttatus sont absentes. ⇒ Herichthys deppii

(L’espèce la plus méridionale trouvée endémique aux rivières Nautla et Misantla dans l’État de Veracruz) .
1 ». Nageoire anale avec cinq épines modales. ⇒ 2
2 .Tête grosse, longue et pointue, généralement plus longue (toujours supérieure à 90 %) que la profondeur du corps au-dessus de la base pelvienne ;
Bouche très grande, mâchoire inférieure dépassant devant la mâchoire supérieure ;
Corps long et étroit. ⇒ Herichthys steindachneri

(Herichthys steindachneri endémique du Rio Gallinas, un affluent isolé du Rio Panuco)
2′. Tête plus courte, longueur généralement bien inférieure à la profondeur du corps au-dessus de la base pelvienne ;
Mâchoires égales ou supérieures légèrement saillantes ;
Bouche petite. ⇒ 3
3 . Spécimens non reproducteurs avec joues et lèvres bleues mais corps sans marques bleues ;
Coloration nuptiale jaune-noir au lieu de blanc-noir;
Dents pharyngées le long de la ligne médiane de la mâchoire pharyngée inférieure (LPJ) fortement molariformes dans tous les spécimens. ⇒ Herichthys labridens

(Espèce endémiques aux lagons de la région de Laguna de la Media Luna (San Luis Potosl) à des altitudes comprises entre 1 000 et 1 100 mètres d’altitude) .
3′. Spécimens non reproducteurs avec des taches bleues opalescentes sur le corps ;
Spécimens reproducteurs avec une coloration noire sur la tête limitée au-dessous d’une ligne partant de l’angle de la bouche. ⇒ 4
3 ». Spécimens non reproducteurs sans taches bleues opalescentes sur le corps ;
Spécimens reproducteurs avec une coloration noire sur la tête atteignant le haut et touchant le bord inférieur de l’œil. ⇒ 7
Dents de la mâchoire buccale pointues ;
Femelles reproductrices entièrement blanches, mâles reproducteurs entièrement noirs ;
Tête relativement longue (36 % en SL contre 33 % – 34 %) ;
Museau : 37 % contre 30 % – 35 % ;
Distance prédorsale : 36 % contre 32 % – 33 % ;
Nageoire dorsale relativement courte : 64 % contre 69 % – 75 % ;
Base de la nageoire dorsale : 49 % contre 54 % – 56 % ;
Epines de la nageoire dorsale : 51 % contre 54 – 58 % ;
Nageoire pectorale plus courte : 24 % contre 25% – 27% ;
Plus d’écailles sur la ligne latérale : 33 contre 27 – 31 ;
Polymorphe en forme de tête, robustesse LPJ et dentition. ⇒ Herichthys minckleyi
(Espèce endémique du bassin désertique endoréique de Cuatro Cienegas, Coahuila, une partie isolée du bassin du Rio Bravo, avec Herichthys cyanoguttatus, l’espèce la plus septentrionale) .
4′. Dents de la mâchoire buccale tronquées, en forme d’incisives ;
Femelles reproductrices avec une quantité variable de noir sur la moitié inférieure de la tête et du corps ;
Tête relativement courte : 33 % – 34 % dans SL contre 36 % ;
Museau : 30 % – 35 % contre 37 % ;
Distance prédorsale : 32 % – 33 % contre 36 % ;
Nageoire dorsale relativement longue ; 69 % – 75 % contre 64 % ;
Base de la nageoire dorsale : 54 % – 56 % contre 49 % ;
Epines de la nageoire dorsale : 54 – 58 % contre 51 % ;
Nageoire pectorale plus longue : 25 % – 27% contre 24% ;
Moins d’écailles sur la ligne latérale : 27 – 31 contre 33. ⇒ 5
5 . Rayons des nageoires pectorales : 13 ;
Taches opalescentes sur le corps et la tête très petites et blanchâtres ;
Coloration nuptiale noire sur la tête très limitée, préopercule entièrement blanc. ⇒ Herichthys tamasopoensis

(Espèce endémique du Rio Gallinas, un affluent isolé du Rio Panuco).
5′. Rayons des nageoires pectorales 14. ⇒ 6
Taches opalescentes sur le corps et la tête, petites ;
Couleur nuptiale noire très limitée sur la tête, préopercule et toute la zone du dessous de la bouche est blanche. ⇒ Herichthys cyanoguttatus
(L’espèce atteignant l’extrême nord, répartie entre les aires de répartition de Herichthys minckleyi au nord-ouest et de Herichthys carpintis au sud depuis le Rio Bravo et le Rio Nueces au Texas jusqu’au sud, y compris les bassins de San Fernando et Soto la Marina) .
6′. Taches opalescentes sur le corps et la tête grandes ;
Couleur noire de reproduction sur la tête la plus développée parmi le groupe Herichthys cyanoguttatus atteignant jusqu’à l’angle de la bouche avec toute la partie inférieure de la tête noire. ⇒ Herichthys carpintis

(Espèce distribuée entre Herichthys cyanoguttatus au N et Herichthys tepehua au S dans tout le bassin de Panuco, à l’exception de son cours supérieur (où il a été introduit artificiellement).
6 ». Taches opalescentes si agrandies qu’elles fusionnent en une coloration corporelle unique aigue-marine-turquoise ;
Certains spécimens avec des points orange/brun rouille sur le visage et les joues (sinon présents uniquement chez Herichthys deppii, où ils sont beaucoup mieux développés et plus grands) ;
Quantité de coloration noire sur la tête intermédiaire entre Herichthys carpintis et Herichthys cyanoguttatus. ⇒ Herichthys tepehua

(Espèce distributaire entre Herichthys carpintis au N et Herichthys deppii au S dans les systèmes fluviaux Tuxpan/Pantepec, Cazones, Tecolutla, Tenixtepec et Solteros) .
- 7. Marques rouges sur les nageoires non appariées et points rouges sur le corps ;
Tache rouge (contre violette/magenta) à l’aisselle de la nageoire pectorale ;
Reproduction coloration avec un fond blanc jaunâtre; développement variable des dents molariformes LPJ ;
Espèce dimorphe avec des formes lacustres et fluviales distinctes en termes de coloration et de forme corporelle. ⇒ Herichthys pantostictus

(Forme lacustre ; seules les espèces répandues du groupe d’espèces Herichthys labridens sont endémiques à pratiquement tout le bassin fluvial Tamesi-Panuco, y compris le Rio El Salto, mais à l’exception des zones de haute altitude du Rio Verde, du Rio Gallinas et du cours supérieur du Rio Moctezuma).
7′. Aucune marque rouge sur les nageoires non appariées et sur le corps ;
Une tache violette/magenta (contre rouge) à l’aisselle de la nageoire pectorale ;
Coloration reproductrice avec un fond blanc pur;
Dents LPJ molariformes invariablement bien développées. ⇒ Herichthys pame


–
NOMS
NOM COMMUNS
- Cichlidé de MINCKLEY ;
- Cichlidé de Cuatro Ciénegas ;
- Mojarra de Cuatro Ciénegas ;
- …
–
SYNONYMES
https://www.desertfishes.org/dfc/obits/wlm/wlm_obit.html
- Cichlasoma minckleyi.
–
ETHYMOLOGIE
Le nom spécifique de cette espèce a été choisi pour rendre hommage à l’ichtyologue Wendell L. MINCKLEY (1935–2001) de l’Arizona State University qui a étudié l’écologie de Cuatro Ciénegas.
Wendel Lee MINCKLEY (1935 – 2001), connu parmi ses collègues et étudiants comme « Minck » était Ichtyologiste et biologiste de la conservation spécialisé dans l’étude de Cuatro Ciénegas qui est l’unique zone marécageuse dans le désert.
Wendel Lee MINCKLEY était aussi un contemporain d’un autre Ichtyologiste, Robert Rush MILLER, avec lequel il a écrit « Freshwater Fishes of México », un ouvrage standard sur l’ichtyologie mexicaine, paru 3 ans après sa mort.
–
A propos de FRESHWATER FISHES OF MEXICO…
Freshwater Fishes of México est un ouvrage de référence et certainement l’œuvre majeure de Robert Rush MILLER.

Il était l’un des plus grands spécialistes mondiaux des poissons d’eau douce, a été professeur émérite au département de zoologie et conservateur au musée de zoologie de l’université du Michigan de 1948 à sa retraite en 1987.
Pour la rédaction de cet ouvrage, MILLER s’est entouré de :
- L. Minckley (1935-2001), professeur émérite de biologie à l’université d’État de l’Arizona ;
Steven Mark NORRIS, un ichtyologue qui enseigne la biologie à l’université d’État de Californie Channel Islands.
Très longtemps attendu par les scientifiques, il a été accueilli avec enthousiasme non seulement par les spécialistes des poissons mexicains, mais aussi par tous les ichtyologues travaillant en Amérique centrale et du Nord.
Ce livre a aussi trouvé également un écho auprès des aquariophiles amateurs, des gestionnaires de pêche, des biologistes de la conservation et des aménageurs et gestionnaires du territoire.
Bien que le Mexique ne représente qu’un cinquième de la superficie des États-Unis continentaux, il abrite près des deux tiers d’espèces de poissons d’eau douce en nombre égal à celui des eaux des États-Unis et du Canada réunis.
Cette riche biodiversité s’explique par la grande variété de sa géographie physique, son étendue en latitude, le plus vaste réseau fluvial d’Amérique centrale et, paradoxalement, ses océans : de nombreuses espèces marines ont quitté l’eau salée pour les sources intérieures et n’y sont jamais retournées.
Nul ne connaissait mieux ces eaux douces mexicaines et les poissons qui les peuplent que le regretté Robert Rush MILLER, ichtyologue de renommée mondiale.
Pionnier dans son domaine, MILLER entreprit sa première expédition de terrain il y a plus de cinquante ans et, au cours des décennies suivantes, rassembla les informations nécessaires à la rédaction de la première encyclopédie de l’ichtyologie des eaux douces du Mexique.
Offrant des clés d’identification pour plus de 500 espèces indigènes, accompagnées de cartes de répartition détaillées et d’illustrations.
Ce livre propose un panorama historique de l’ichtyologie du pays, ainsi qu’une synthèse de la biogéographie unique des poissons mexicains et de leur statut de conservation actuel.
Organisées par famille, les fiches descriptives des espèces sont complétées par des galeries en couleurs présentant des photographies de poissons vivants dans leurs milieux et habitats naturels.
Explorant les enjeux écologiques, biologiques et taxonomiques, l’ouvrage aborde également l’histoire évolutive de l’ichtyofaune elle-même et l’histoire des scientifiques qui l’ont étudiée au cours des derniers siècles.
–
BIOGRAPHIE
Professeur émérite au Département de biologie de l’Arizona State University, Wendell MINCKLEY était mieux connu sous le nom de « Minck » par ses collègues et amis et par des générations d’étudiants diplômés, est décédé le 22 juin 2001 à « Désert Hôpital Samaritan » à Mesa en Arizona, suite à des complications associées à traitement du cancer.

Il avait 65 ans et vivait à Tempe.
Le Dr MINCKLEY est venu en Arizona State University en 1963, où il a mené des recherches et enseigné pendant 38 ans, enseignant même après sa retraite en 2000 pour devenir professeur émérite.
Wendell Lee MINCKLEY est né le 13 novembre 1935 à Ottawa, Kansas.
Il était diplômé de la Kansas State University en 1957, où il s’était spécialisé en Biologie de la faune et des pêches.
Non loin de là, à l’Université du Kansas, il a obtenu un master en zoologie avec une spécialisation en ichtyologie en 1959.
Il a obtenu un doctorat en biologie à l’Université de Louisville en 1962, où il étudia avec le professeur Louis KRUMHOLTZ.
Sa thèse était exhaustive étude de l’écologie aquatique de Doe Run, un cours d’eau du comté de Meade, Kentucky.
Son domaine de prédilection fut la géologie, une discipline qui allait s’avérer fondamentale pour ses études ultérieures sur la zoogéographie des poissons du Sud-Ouest.
Un an après sa nomination au département de biologie de l’Université Western Michigan il a rejoint l’Arizona State University en 1963 en tant que professeur assistant de zoologie.
Le département de zoologie était nouveau (1962), occupait une nouvelle aile (1963) et se formait ses premiers étudiants diplômés.
Il obtient son doctorat en zoologie en 1965.

Le professeur MINCKLEY fut l’un des premiers membres du corps professoral recrutés pour déplacer la université depuis ses racines traditionnelles sous le nom d’ « Arizona Territorial Normal School » vers l’Université de Recherche telle qu’elle est aujourd’hui.
Il a joué un rôle fondamental dans cette transformation alors qu’il remportait d’importants prix de recherche, formait des étudiants, et a mis en place un programme de recherche reconnu internationalement en écologie aquatique, Ichtyologie systématique et biologie de la conservation.
À l’ASU, le Dr MINCKLEY et ses étudiants étudiaient les sciences aquatiques écosystèmes et poissons du Sud-Ouest.
Il fut l’auteur et éditeur de trois livres ainsi qu’environ 175 articles de revues et chapitres de livres.
En 1973, il a publié « Fishes of Arizona », le premier compendium des poissons de cette région.
En équipe avec ses collègues, il a rédigé d’importants articles sur :
- La rivière Gila et sur le fleuve Colorado (1972, 1976-79) ;
- Les poissons du Rio Yaqui dans le nord-ouest du Mexique (1980) ;
- La communautés végétales et animales aquatiques du sud-ouest (1982, 1984) ;
- Le changement environnemental dans les cours d’eau de l’Arizona du XIXe siècle à la fin du XXe siècle (1985) ;
- Le Zoogéographie des poissons d’eau douce de l’ouest du Mexique et des États-Unis (1986).
Il est un contributeur majeur à la prochaine publication de « Fishes of Mexico. »
Les recherches du Dr MINCKLEY ont documenté les changements d’habitats et Organismes.
La demande humaine croissante d’eau dans le sud-ouest américain a commencé vers 1900 et continue encore aujourd’hui, perturbant les habitats aquatiques et poussant toutes sauf une poignée d’espèces de poissons indigènes à l’extinction, ou au bord du extinction.
En 1968, le Dr MINCKLEY Fut l’un des fondateurs du « Desert Fishes Council », un groupe dédié à la conservation des habitats aquatiques et des poissons dans les zones arides Terres.

La mission du Desert Fishes Council (DFC) est de préserver l’intégrité biologique des écosystèmes aquatiques désertiques et des formes de vie qui y sont associées, d’organiser des colloques pour présenter les résultats de recherches et les initiatives de gestion menées dans ce domaine, et d’assurer une diffusion rapide des informations relatives aux activités du Conseil et de ses membres.
Il était infatigable dans ses efforts de conservation, combinant recherche et formation et un service auprès des agences étatiques et fédérales.
En dehors de son aspect technique, des articles scientifiques, il écrivait des articles pour éduquer le grand public parce qu’il pensait que ces articles sont des vecteurs importants pour promouvoir une éthique de la conservation.
Il était un naturaliste talentueux, qui lui a bien servi en tant que biologiste de la conservation.
Cinq espèces découvertes par ses soins (un escargot, un scorpion, un scarabée, une mouche et un cichlidé) se voient attribuer le Nom de « Minckleyi » en reconnaissance de ses découvertes.
Plusieurs de ces espèces sont de Cuatro Cienegas, Coahuila, Mexique.
Le professeur MINCKLEY a joué un rôle clé dans cette région étant désignée comme zone naturellement protégée afin de préserver sa Flore et faune remarquables.
Environ 45 étudiants ont obtenu des diplômes de doctorat et de master sous la direction du Dr MINCKLEY.
En 1990, le Graduate College de l’ASU lui a décerné une récompense sa plus haute distinction : Enseignant exceptionnel des étudiants diplômés.
Le Professeur MINCKLEY a également reçu des prix au-delà de son université d’origine :
- Le prix de Excellence de l’American Fisheries Society (1980) ;
![]()
- Département de l’Intérieur des États-Unis avec Mention élogieuse pour service (1987) ;
- Le Certificat de mérite scientifique de l’American Association pour l’avancement des sciences, division du Sud-Ouest et des Rocheuses (1988) ;
- Le Certificat de mérite en sciences biologiques de l’Universidad Autónoma de Baja California Norte (1990) ;
- Le Certificat d’appréciation pour service exceptionnel du Conseil national de la recherche (1991).
Il a été finaliste pour un « Pew Scholars Award » en Conservation et environnement (1992).
Tout au long de sa carrière, le Dr MINCKLEY a servi au niveau étatique et fédéral ainsi que des sociétés professionnelles. Protection de la rare aquatique les ressources du Sud-Ouest ont toujours été primordiales.
Ses opinions pourraient être Controversé car ils variaient fréquemment selon les recommandations des agences et les développeurs concernant l’utilisation et la protection des ressources aquatiques.
Il a servi de nombreux comités d’État et fédéraux chargés d’examiner le statut des poissons en danger espèces et leurs habitats.
De 1984 à 1985, il a été biologiste principal des pêches à Centre national de la Pisciculture et de la Technologie Dexter, une installation fédérale au Nouveau-Mexique dédiée à la conservation des espèces en danger.
Plus récemment, il a servi au service des États-Unis, au Comité de l’Académie des sciences, chargé d’examiner le suivi et la recherche de Glen Canyon Programme.
Le Dr MINCKLEY était une autorité internationalement respectée dans le domaine la systématique, l’écologie et la biologie de la conservation des poissons de la Sud-ouest des États-Unis et Mexique.
Ces animaux l’intéressaient comme étant exceptionnels exemples du processus évolutif, et ses recherches furent parmi les premières à attirer l’attention sur les forces qui ont conduit cette faune à l’extinction.
Avec James DEACON, il a publié « Battle Against Extinction » (1990), un livre qui se lit à des temps comme une élégie pour une faune disparue.


Dans la préface, ils écrivaient :
« Les conservationnistes sont d’accord en tant que groupe… que la diversité doit être maintenue pour le bien-être de la biosphère, ainsi que le bien-être humain.
Ceux qui Embrassent d’autres philosophies est souvent tout aussi fermement convaincu de leur des vues alternatives, et uniquement une éducation fondée sur des données tangibles et logiques, des résultats documentables de la recherche et de l’observation peuvent être attendus pour modifier leur Esprits ».
Ce volume fournit de telles informations, et nous espérons qu’il sera largement utilisé comme référence pour fournir des exemples de ce qui a été appris et accompli dans traiter avec un groupe obscur d’animaux qui dépendent de l’eau dans un cas improbable endroit. »
Les écosystèmes aquatiques ont offert au professeur MINCKLEY la Contexte pour comprendre l’évolution des poissons.
Il a passé sa carrière à combattre contre les changements anthropiques détruisant les organismes qu’il aimait et leur habitats.
Ailleurs dans la bataille contre l’extinction, Robert et Frances MILLER avec Clark HUBBS, cofondateurs du « Désert Fishes Council », ont observé :
« Il existe de solides raisons biologiques de conserver des échantillons de pays vierges dans perpétuité.
Avec le temps, la majeure partie du désert sera utilisée par les humains à une seule raison ou autre irrigation, pâturage, exploitation minière ou aires de jeux mais inévitablement, les scientifiques devront connaître la situation initiale.
Un contrôle est une partie de base de chaque expérience.
La conservation de la nature sauvage pour l’entretien des maladies biologiques la diversité est une nécessité pour la survie des humains eux-mêmes.
Ce que nous devons avoir est une Terre domestiquée et homogène ;
Pour beaucoup, ce serait bien moins un endroit fascinant où vivre. »
Le professeur MINCKLEY a consacré sa vie professionnelle pour éviter ce résultat.
Il préférait une terre riche en diversité végétale et des animaux qui l’ont tant fasciné tout au long de sa vie.
–
DESCRIPTION
Herichthys minckleyi est une espèce unique car elle présente deux morphotypes distincts :
- Un morphotype trapu ;
- Un morphotype élancé.
Les morphotypes élancés ont généralement un intestin plus long et se nourrissent de petits poissons.
Le morphotype trapu, en revanche, a un intestin plus court et se nourrit de gastéropodes.
Ce poisson est également connu pour ses deux morphotypes trophiques distincts Þ la dentition des individus varie au sein d’une même espèce en fonction de leurs habitudes alimentaires, indépendamment de leur morphotype.
Les premières études sur la faune des cichlidés de la vallée (LABOUNTY, 1974) ont suggéré que l’ampleur des différences biologiques entre les trois morphes vivant dans les sources thermales montrait l’existence d’au moins trois espèces différentes.
Ces deux morphotypes sont le morphotype « à petites dents » et le morphotype « à grandes dents ».
- Le morphotype « à petites dents» possède des dents pharyngiennes papilliformes, c’est-à-dire que les dents présentes sur une arcade dans la gorge du poisson sont petites, pointues et en forme de crayon.
- Les poissons du morphotype « à grandes dents» possèdent de grandes dents pharyngiennes molariformes plates.
Certaines études mentionnent l’existence d’une troisième forme, caractérisée par une dentition pharyngienne intermédiaire entre les formes molariforme et papillaire.
Toutefois, cette forme représente moins de cinq pour cent de la population totale.
La différence entre ces formes constitue également un critère de distinction important entre Herichthys minckleyi et les autres membres du groupe Herichthys.
–
Phylogénie
On pensait auparavant que la divergence dentaire était la preuve de deux espèces distinctes ou d’une hybridation.
Cependant, les recherches génétiques montrent que les morphotypes appartiennent toujours à la même espèce et ne proviennent pas de lignées génétiquement distinctes.
Néanmoins, les polymorphismes liés aux ressources peuvent être l’étape initiale de la spéciation et de la divergence des populations, et la région de Cuatro Ciénegas et cette espèce ont été comparées aux îles Galápagos et aux pinsons de Darwin.
–
MORPHOLOGIE
L’EVOLUTION
Des études génétiques ont révélés que la variation génétique entre les trois formes ne correspondent pas.
La variation était néanmoins trop petite pour garantir une nouvelle espèce.
Surpris, les scientifiques ont décidé qu’une nouvelle espèce s’est formée dans un entourage homogène et constant, ceci néanmoins sans produire assez de variations génétiques.
Ils ont également décidé que les différences entre les trois formes, selon le standard générale taxonomique, sont assez grandes pour les classer comme trois espèces différentes.
Néanmoins, ils ont considéré comme possible mais improbable que les trois types de Herichthys Minckleyi fassent partie d’une sorte polymorphique.
Une situation comme celle-ci ne s’est pas encore produite.
Des observations de KORNFELD ont révélé à partir d’une population échantillon de 39 couples que plus de 50% s’étaient formé entre deux formes différentes.
Ils ont constatés que les couples mixtes produisent des jeunes qui, individuellement possèdent des caractéristiques de l’un des parents. Le nombre de jeunes ayant des caractéristiques mixtes était représenté dans un pourcentage si petit qu’il était considéré sans importance.
Selon SAGE et SELANDER, Herichthys Minckleyi descend du Herichthys Cyanoguttatus.
Ils ont suggérés que son origine venait d’un cichlidé avec une denture papilliforme, emprisonné dans la vallée ou on trouvait un excès d’escargots.
Il viendrait de la vallée que l’on peut atteindre par un petit Canyon : la vallée Ocampo. Dans cette vallée on trouve quelques sources d’eau.
Ces sources montrent des caractéristiques ressemblant à celles du Rio Salado, se situant à l’est de la vallée de Cuatro Cienegas et ressemblant aussi à la partie inférieure du vieux bassin de Cuatro Cienegas.
Dans les sources (Ocampo) on trouve Herichthys Cyanoguttatus, ensemble avec Astyanax fasciatus et Gambusia Marshi.
Ces sortes font également partie de la faune de Rio Salado.
Les deux dernières espèces se trouvent également dans le Cuatro Cienegas, mais dans une autre forme géographique.
L’ancêtre de Herichthys minckleyi a donc pu s’isoler au milieu de la vallée, dans un entourage qui se différencie assez beaucoup de celui du bassin Rio Salado, à cause de quoi l’évolution de la population isolée prend garanti une autre direction que celle de la population de Rio Salado.
On suggère aussi que ce cichlidé se trouve encore dans un stade tôt de formation/transformation vers une nouvelle espèce et que dans le future les trois types vont de plus en plus choisir des partenaires de leurs même type à cause de quoi une nouvelle espèce se formera.
–
CARACTERISTIQUES
L’espèce est unique car elle possède plusieurs formes dans la nature, spécialisées pour différentes habitudes alimentaires. Un type présente de petites dents pour se nourrir principalement de matières végétales, tandis que l’autre type présente des dents plus grandes et une bouche plus large, parfaites pour écraser les escargots.
Ils semblent occuper le même territoire et constituent en fait une seule population de poissons !
Trois formes différentes existent de ce poisson.
Les formes se différencient en ce qui concerne la morphologie, la longueur du canal digestif et la denture pharyngale et enfin par le régime alimentaire.
La forme de la tête de chaque morphotype est très caractéristique et permet leur identification.
De plus, la tête du morphotype molluscivore se distingue des deux autres morphotypes, vue de dessus ou de face, par ses muscles massifs qui soutiennent une mâchoire pharyngienne plus large, lui donnant ainsi une apparence nettement plus imposante.
–
LES TYPES D’HERICHTHYS
Le classement par « différences de formes » des Herichthys minckleyi n’est pas la méthode qu’on retenu les ichtyologues, qui ont pris comme critère de sélection la dentition de ces poissons pour les classer.
Toutes les observations faites sur la physiologie de ce poisson confirment qu’Herichthys minckleyi est une espèce polymorphe Þ une espèce qui peut se présenter sous diverses formes.


Cette variation se produit lorsque les sous-groupes deviennent isolés et se développent indépendamment ou en raison de la pression concurrentielle.
Ce dernier est particulièrement évident pendant la pénurie alimentaire.
Ensuite, pour certains individus, les sources alimentaires secondaires peuvent devenir de plus en plus primaires.
Un phénotype légèrement différent peut soudainement être utile et une zone de recherche de nourriture partagée garantit alors que les animaux du même type s’accouplent les uns avec les autres.
De cette façon, les individus séparés forment un groupe séparé et lorsque ces groupes ont la possibilité de se développer suffisamment longtemps et suffisamment loin, de nouvelles espèces peuvent apparaître de cette manière.
Mais c’est Herichthys minckleyi pas encore.
Au moins, selon les vues généralement acceptées.
On distingue cependant trois types principaux :

Plaques dentaires pharyngiennes inférieures d’Herichthys minckleyi.
(a) Les papilliformes sont caractérisés par de petites dents pointues et une morphologie pharyngienne légère.
(b), (c) La présence de dents pharyngiennes élargies et aplaties a permis de classer Herichthys minckleyi parmi les molariformes.
Bien que le nombre de dents augmente généralement avec la taille, on observe une variation importante chez les molariformes.
Certains molariformes présentent un nombre important de dents molariformes sur leur mâchoire pharyngienne, tandis que d’autres, de taille similaire, peuvent en présenter un nombre bien moindre.
Chez les individus plus petits (non représentés), de longueur < 50 millimètres, la morphologie pharyngienne est difficile à déterminer avec certitude.
Tout le monde avec une apparence différente, une habitude et un comportement alimentaires.
Des types intermédiaires (poissons à caractères mixtes) existent toujours entre ces types principaux.
L’ancêtre de toutes ces manifestations est très probablement Herichthys cyanoguttatus, qui dans le passé a pu atteindre la vallée via une connexion temporaire.
Les spécialisations mentionnées s’expriment principalement lors de la pénurie alimentaire mais, quoiqu’il arrive, ces animaux ont conservé leur préférence pour les insectes aquatiques et les larves.
Cela devient clair lorsqu’ils sont conservés dans un aquarium et qu’un menu varié leur est proposé.
Dès que les mangeurs d’escargots peuvent obtenir quelque chose de mieux que des escargots, à savoir quelque chose qui prend moins d’énergie à saisir, casser et macher et ou une valeur nutritive plus élevée, ils ignorent rapidement les escargots.
La proportion d’escargots dans le menu d’un molluscivore varie, selon la saison ou selon la disponibilité de la nourriture.
Les animaux utilisent donc leur spécialisation surtout en période de pénurie, mais dans les zones désertiques, c’est bien sûr généralement la plus grande partie de l’année.
Les caractéristiques qui séparent ces trois types ont une composante héréditaire et ne sont pas seulement le résultat de la plasticité phénotypique (variation due à des facteurs environnementaux), mais ces caractéristiques sont déclenchées par des facteurs environnementaux.
Par exemple, un molluscivore est plus susceptible de développer des dents en forme de molaire lorsqu’il est confronté à des aliments durs.
À l’inverse, cependant, il n’est pas vrai que les dents en forme de molaire disparaissent également une fois que les animaux ne reçoivent que de la nourriture molle.
Un autre fait qui montre la composante héréditaire est qu’un détracteur ne développera jamais de dents en forme de molaire, même si nous les mettons au régime des escargots.
Le poisson n’a tout simplement pas les bons gènes pour développer des dents en forme de molaire.
1ère forme – Type 1: Détritivores.
 La première forme qui est la plus répandue et le plus primitive est le détritivore, il se nourrit essentiellement de détritus.
La première forme qui est la plus répandue et le plus primitive est le détritivore, il se nourrit essentiellement de détritus.
Ce type se trouve le plus souvent dans les champs Nymphéa.
La première forme est la forme la plus abondante, elle est également considérée comme la plus primitive et ancestrale par rapport aux deux autres.
Les poissons de cette forme possèdent un corps haut et une bouche papiliphormique garnie de fines dents pharyngées façonnées pour désintégrer les aliments mous.
La forme de poisson est la plus courante rencontrée et aussi la plus largement distribuée.
Les poissons de cette forme possèdent un corps assez grand et des dents sous-développées.
Cet Herichthys minckleyi a une mâchoire pharyngale avec des petites dents coniques (papilliformes)qui sont employées pour broyer de la nourriture douce.
Ces poissons ont des dents au niveau de la gorge qui sont coniques Þ On parle de dents « papilliformes ».
Les dents de la gorge de ce type sont petites et rapprochées.
Ces types de dents conviennent aux petits aliments mous tels que les détritus, les larves d’insectes et les algues. Pour la partie végétale, ces animaux sont équipés d’un système intestinal plus long.
La mâchoire a également une légère toile de muscle, on dit même que les muscles de la mâchoire sont faibles.
Ce type de prothèse se prête bien à l’ingestion de petits aliments mous tels que les détritus, les larves d’insectes et les algues car, en effet, la principale source de nourriture des poissons de cette forme est constituée de détritus.
Ce type de poisson se rencontre le plus souvent dans les champs de Nymphéa et pour digérer des aliments d’origine végétale, ces animaux disposent d’un système intestinal plus long.
Il faut bien savoir que, par défaut, tous les types (3) commencent d’abord par se nourrir à base de détritus, ce n’est que plus tard, lors de leur croissance, que la spécialisation alimentaire intervient et se manifeste.
Ce type est donc aussi considéré comme le plus original, et il est similaire à l’omnivore Herichthys cyanoguttatus.
Ces animaux ont (surtout en dehors de la saison de reproduction) tendance à former des groupes : Les formes détritivores, et dans une moindre mesure les formes molluscivores, sont des poissons grégaires.
On peut les observer en groupes, partageant un vaste territoire.
Ils constituent des territoires généralement situés proches des rives des sources, délimités par des éléments tels que le bord de la source, un rocher ou un amas de végétation.
Leur superficie peut atteindre environ 3 mètres carrés.
Ces territoires sont habités par un grand nombre de poissons, la plupart ignorés par le mâle dominant, généralement de la forme détritivore.
Ce mâle dominant se distingue des autres cichlidés par une bosse nucale presque toujours présente.
De plus, il arbore une coloration dominante Þ un assombrissement de la couleur de base du corps, virant au bleu ou au gris très foncé, presque noir, et des taches bleues aux reflets irisés sur les flancs et les nageoires.
Les mâles patrouillent cycliquement le périmètre de leur territoire, puis se retirent à l’arrière.
Ces territoires abritent une population importante de poissons, majoritairement des cichlidés femelles ou des mâles non dominants de la forme détritivore.

Ces poissons se nourrissent à la surface des feuilles immergées de Nymphea sp. ou des rochers, des détritus, ou encore sous la végétation surplombante.
–
2éme forme – Type 2: Molluscivores
C’est une forme moins abondante que la première et les Herichthys minckleyi évoluent le plus souvent dans des sortes de piscines thermales aux dimensions limitées et situées à proximité de la point de la Sierra San Marcos.

Jeune mâle adulte Herichthys minckleyi -Forme molluscivore.
Ce type de poissons possède un corps élevé et une forte molariphormie composée de dents pharyngées qui sont utilisées pour broyer les coquilles dures des mollusques, tels que les Mexipyrgus endémiques.
Les Herichthys minckleyi de cette forme et de type molluscivores ont des grandes dents pharyngiennes en forme de meule : On parle alors de dents « molariformes ».

Ces éléments leur permettent de casser les coquilles d’escargots.
Cela pourrait s’expliquer par que fait que ces poissons, à l’origine omnivores, se sont retrouvés dans un biotope où les escargots étaient abondants et qu’ils ont commencé à puiser dans cette source de nourriture ce qui leur permis de développer leurs particularités physiques.
Avec des dents de gorge légèrement plus grandes et placées au niveau de la gorge, il est devenu possible à ces poissons de casser les plus petites coquilles d’escargot, ce qui a lancé une ligne de développement vers les dents actuelles en forme de meule.
Ce poisson emploie sa forte mâchoire pour écraser des coquilles de l’escargot endémique (de la famille Mexipyrgus).
Les cichlidés molluscivores, seuls ou en petits groupes, évoluent près du fond détritique des sources à la recherche de leur principale nourriture : les escargots, qu’ils repèrent probablement grâce à leur ouïe.
Ces cichlidés présentent une particularité intéressante : les individus qui chassent les escargots, seuls ou à la tête d’un petit groupe de deux ou trois, arborent parfois une coloration sombre, d’un noir velouté, bien plus foncée que celle des mâles dominants.
Dans cette coloration, les taches bleues irisées scintillent intensément.
Les cichlidés molluscivores se déplacent très près du fond où, de temps à autre, ils orientent leur corps presque perpendiculairement à celui-ci, enfouissant la moitié de leur corps dans le substrat et prenant une bouchée de détritus.
Ils manipulent les détritus dans leur bouche un instant, puis libèrent les plus petits morceaux par leurs branchies.
Les petits escargots sont broyés par leurs mâchoires pharyngiennes et avalés. Les coquilles broyées restantes et les particules plus grosses sont expulsées par la bouche.
Ce comportement est intéressant car il est très similaire à celui d’Herichthys labridens dans les sources de la vallée du Rio Verde, dans le bassin de Panuco, où ces poissons présentent exactement la même coloration.
Cette forme de Herichthys Minckleyi possède en plus un canal digestif qui mesure 25 à 50% de moins que celui du détritivore. On pense que ce type est une évolution du type détritivore.
Avec le temps, la progéniture de ces poissons est devenu aussi de plus en plus capable d’utiliser cette source de nourriture.
Ce type d’Herichthys minckleyi est le plus courant dans les endroits peuplés d’une forte densité d’escargots et, ils aussi la particularité d’être plus facilement identifiables grâce à une musculature dentaire mieux développée ce qui leur confère souvent aussi une tête plus large.
Par conséquent, pour broyer les coquilles de ces mollusque, les muscles de la mâchoire sont forts.
Aussi la longueur de l’intestin représente environ 25 à 50 % de celle de forme papiliphorme.
Sans rapport avec leur morphologie, les poissons de ce type ont nettement plus tendant à former des groupes.
–
3éme forme – Type 3: Piscivores
La troisième forme de poisson est assez rare et possède un corps allongé, c’est le propre de ce type de poisson qui peut nager rapidement et chasser de petits poissons.

Le troisième type d’Herichthys minckleyi peut atteindre une longueur de 15 à 20 centimètres, c’est le moins répandu et c’est la forme piscivore ou indéterminée : Il se nourrit surtout d’Astyanax fasciatus.
Les poissons piscivores adoptent une coloration cryptique à bandes verticales qui favorise leur comportement de prédation.

Cette forme rare n’apparaît que dans quelques sources thermales au pied du San Marcus Y Pinos, une crête montagneuse qui traverse la vallée des Cuatro Ciénegas par le sud.

San Marcus Y Pinos.
Son biotope se situe dans quelques sources au sommet de la crête de San Marcos.
Cette forme se nourrit individuellement et présente un comportement unique.

Forme piscivore.
Lorsqu’elle chasse, sa coloration se transforme en un motif de camouflage brun foncé, jaune, noir et blanc.
Le poisson se repose ensuite au fond des sources, parmi les rochers.
Le motif de coloration du poisson, combiné aux ombres projetées par le soleil sur les vagues, lui offre un camouflage parfait.
Il attend qu’un petit poisson passe près de sa tête.
Le plus souvent, sa proie de prédilection est un Astyanax fasciatus.
Dès que la proie s’approche suffisamment, le prédateur à l’affût surgit du fond pour tenter de l’engloutir, avec un succès mitigé.
Cette troisième forme a une forme corporelle allongée, qui permet une nage plus rapide, cruciale pour attraper des proies.
Ces poissons se caractérisent par un corps plus long et profilé.
Le corps semblable à celui d’un brochet leur permet d’attraper des poissons.
Le tube intestinal de ces animaux est le plus court des trois types d’Herichthys.
Contrairement aux détrivores et molluscivores, les piscivores vivent solitaires.
Bien qu’ils reposent patiemment au fond, le jeu d’ombres, causé par la ride de la surface de l’eau, rend ces Herichthys presque invisibles : Ces poissons se parent de couleurs de camouflage pendant la chasse.
Alors qu’ils reposent patiemment sur le fond, le jeu d’ombres provoqué par l’ondulation de la surface de l’eau rend ces Herichthys pratiquement invisibles.
Cette forme caractérise les Herichthys minckleyi piscivores qui chassent de petites espèces de poissons, principalement des Axtyanax mexicanus.
Cette forme possède à la fois des dents molariphormiques et papiliphormiques et un corps plus allongé qui l’autorise à nager plus vite, ceci pour arriver à se procurer sa plus grande source de nourriture, notamment des autres poissons.
Chez le type piscivore on trouve aussi bien une denture identique au détritivore qu’une denture identique à celle du molluscivore.
Le canal digestif de cette forme est beaucoup plus courte que celui du détritivore et à tendance à ressembler sur celui d’un carnivore.
–
CARACTERES GENERAUX
Corps
Herichthys minckleyi est une espèce unique car elle présente deux morphotypes distincts : un morphotype trapu et un morphotype élancé.
Les individus au corps élancé possèdent généralement un intestin plus long et consomment des poissons plus petits.

À l’inverse, les individus au corps trapu ont un intestin plus court et se nourrissent de gastéropodes.
La différence entre ces formes constitue également une caractéristique distinctive importante entre Herichthys minckleyi et les autres.
Il faut donc considérer qu’il y a trois formes différentes de ces poissons, qui diffèrent principalement par la forme des dents et du corps sachant que de façon générale, Herichthys minckleyi est une espèce polymorphe (une espèce qui peut apparaître sous différentes formes) qui a un corps grand et élancé, légèrement aplati sur les côtés.
La différence entre ces formes constitue également une caractéristique distinctive importante entre Herichthys minckleyi et les autres membres du groupe Herichthys.
Historiquement ces formes étaient autrefois considérées comme des espèces distinctes.
Chacun avec une apparence, une habitude de recherche et un comportement différents.
Parmi ces principaux types, il existe aussi (bien que rarement) des types intermédiaires (poissons aux caractéristiques mixtes).
L’ancêtre de toutes ces manifestations est très probablement Herichthys cyanoguttatus, qui aurait pu atteindre la vallée par une connexion temporaire dans le passé.
⇒ La rareté géographique des ressources alimentaires détermine en partie la spécialisation et, in fine, la morphologie des Herichthys minckleyi
Les spécialisations mentionnées sont principalement exprimées en période de pénurie alimentaire.
Cependant, les animaux ont conservé leur préférence pour les insectes aquatiques et les larves.
Cela devient évident lorsqu’ils sont gardés en aquarium et qu’on leur propose un menu varié.
Dès que les mangeurs d’escargots peuvent avoir quelque chose de mieux, c’est-à-dire quelque chose qui demande moins d’énergie à atteindre et ou a une valeur nutritionnelle plus élevée, ils ignorent rapidement les escargots.
La proportion d’escargots dans le menu d’un molluscivore varie, saisonnière ou autrement, selon la disponibilité alimentaire.
Les animaux utilisent principalement leur spécialité en période de rareté, mais dans les zones désertiques, c’est bien sûr souvent la plus grande partie de l’année.
Les caractéristiques qui distinguent ces trois types les uns des autres ont une composante héréditaire et ne résultent pas uniquement de la plasticité phénotypique (variation due à des facteurs environnementaux).
Cependant, il est vrai que ces caractéristiques sont déclenchées par des facteurs environnementaux.
Par exemple, un mollusquivore est plus susceptible de développer des dents molariformes face à un aliment dur.
Inversement, cependant, il n’est pas vrai que les dents molariformes disparaissent dès que les animaux ne reçoivent que de la nourriture molle.
Un autre fait qui montre la composante héréditaire est qu’un détrivore ne développera jamais de dents molariformes, même si l’on les met sous régime d’escargots. Les poissons manquent tout simplement des bons gènes pour développer des dents molariformes.
La longueur totale de ces poissons est d’environ 18 à 20 centimètres.
Dans un aquarium, les animaux grandissent considérablement.
–
Tête
La différence entre la mâchoire pharyngée papilliforme (supérieure) et molariforme (inférieure) détermine les différents morphes.
La tête est relativement grande, la bouche est terminale et les lèvres sont charnues.

Ce poisson est également connu pour ses deux/trois morphes trophiques distinctes déterminés à partir de la dentition des individus qui varie au sein d’une même espèce en fonction de leurs habitudes alimentaires, quelle que soit leur forme corporelle.
Si ce poisson est bien connu pour ses deux morphotypes distincts, sa dentition varie au sein d’une même espèce en fonction des habitudes alimentaires, indépendamment de la morphologie.
Ces deux morphotypes sont le morphotype à « petites dents » et le morphotype à « grandes dents ».
Le morphotype à « petites dents » possède des dents pharyngiennes papillaires (dents pharyngées papilliformes), c’est-à-dire que ses dents petites, pointues et fines en forme de crayons sont disposées sur une arcade pharyngienne, dans la gorge du poisson.
Les poissons du morphotype à « grandes dents » possèdent de grandes dents pharyngiennes molaires et plates.
Certains auteurs mentionnent un troisième morphotype, présentant une dentition pharyngienne intermédiaire entre molariforme et papilliforme.
Cependant, ce morphotype représente moins de 5 % de la population totale.

La dentition divergente était auparavant considérée comme la preuve de deux espèces distinctes ou d’une hybridation ; cependant, des recherches génétiques montrent que les morphes appartiennent toujours à la même espèce et ne proviennent pas de lignées génétiquement distinctes.
Cependant, les polymorphismes des ressources peuvent être l’étape d’initiation vers la spéciation et la divergence des populations, et la région des Cuatro Ciénegas, cette espèce a été comparée aux îles Galápagos et aux pinsons de DARWIN.
–
Nageoires
Herichthys minckleyi possède une queue arrondie ainsi que ses nageoires pectorales, reliées au corps par un pédoncule caudal fin.
–
TAILLE
Herichthys minckleyi peut atteindre une longueur de 15 à 20 centimètres.
En moyenne, Herichthys minckleyi atteint près de 18 centimètres de long (LS) pour les mâles tandis que les femelles restent légèrement plus petites.
–
COLORATION
Herichthys minckleyi est un cichlidé dont la coloration, sans être marquante, est plutôt attrayante.
La coloration de base des individus non reproducteurs présente des variations allant du gris clair ou beige au jaune verdâtre, en passant par le vert foncé ou le gris.
Le corps est marqué de noir, bien que ces marques varient considérablement d’un individu à l’autre.
Des taches bleues ou jaunâtres sont réparties sur le corps. Herichthys minckleyi présente un fort dichromatisme sexuel en comparaison avec les autres Herichthys.
La coloration noire recouvre l’ensemble du corps et des nageoires chez la plupart des individus.
Un spécimen adulte de la forme détritivore, à coloration albinos (jaune canari vif et yeux rouges), a été découvert par Gary KRATOCHVIL à Poza de la Becerra en 1993 (KONINGS, 1994) : Cela relève de l’exception.
Son apparence peut changer en quelques secondes lors de comportements agressifs, compétitifs ou de reproduction.
Cela signifie que les mâles et les femelles sont colorés différemment, au moins dans la phase de reproduction.
A ce moment, la femelle prend une couleur très claire, à devenir presque blanche tandis que le mâle devient sombre, presque noir.
Habituellement, cela est causé par la sélection sexuelle, mais avec Herichthys minckleyi, c’est un peu différent.
Cela signifie que les mâles et les femelles Herichthys minckleyi sont colorés différemment, du moins lors de la phase de reproduction.
En période normale, la coloration varie du beige au noir, en passant par le gris clair, le vert jaunâtre et le vert foncé.

La couleur varie du beige, gris clair, vert jaunâtre, vert foncé à brun foncé ou noir, avec quatre à sept barres sur la face centrale postérieure du poisson.
Herichthys minckleyi présente quatre à sept barres sur la partie postérieure médiane du corps, généralement sur un léger quadrillage.
Chez les subadultes, une tache peut être présente sur la partie postérieure, mais cette caractéristique disparaît à l’âge adulte.
Les adultes peuvent également développer des mouchetures noires sur le corps et des mouchetures claires sur les nageoires.
Le poisson possède deux lignes latérales, une supérieure et une inférieure, entre lesquelles se situent les quatre à sept barres mentionnées précédemment.
Son apparence peut changer en quelques secondes lors de comportements agressifs, compétitifs ou de reproduction.
https://www.desertfishes.org/dfc/na/cichlida/cichlaso/cminckle/i_cminck.shtml
Sa coloration varie du beige au noir, en passant par le gris clair, le vert jaunâtre et le vert foncé.
Il présente quatre à sept barres sur la partie postérieure médiane du corps, généralement sur un léger quadrillage.
Chez les subadultes, une tache peut être présente sur la partie postérieure, mais cette caractéristique disparaît à l’âge adulte.
Les adultes peuvent également développer des mouchetures noires sur le corps et des mouchetures claires sur les nageoires.

Le poisson possède deux lignes latérales, une supérieure et une inférieure, entre lesquelles se situent les quatre à sept barres mentionnées précédemment.
La couleur sombre des mâles dominants est déclenchée par des interactions compétitives avec leur propre sexe.
Dans la nature, les rivaux se cachent constamment avec l’intention de s’emparer du territoire.
La couleur foncée sert de dissuasion.
Ceci est probablement dû au fait que la dominance est plus facile à établir et à maintenir dans un aquarium.
Les rivaux y sont éliminés rapidement et les mâles subordonnés peinent à accéder à une position sociale plus élevée.
Un mâle dominant acquiert ainsi une sorte de monopole dans l’aquarium, ce qui réduit les besoins de défense.
Dans la nature, de nouveaux rivaux rôdent constamment et imposent une attention constante des mâles dominants.
C’est pourquoi les animaux conservent leurs couleurs sombres d’avertissement.
En aquarium, les mâles deviennent rarement entièrement noirs.
On observe également une différence significative dans le comportement reproducteur du mâle en aquarium : il est beaucoup plus attentionné que dans la nature.
En plus de la discrimination des couleurs, il existe également une différence de longueur.
Les mâles grandissent en moyenne de 10% et portent d’ailleurs souvent une bosse sur le front.

Ces barres reposent généralement sur un léger hachurage croisé.
Lorsque les poissons sont subadultes, une tache peut apparaître à l’arrière du poisson, mais cette caractéristique disparaît à mesure que le poisson atteint l’âge adulte.
Les adultes peuvent également développer des taches noires sur le corps et des taches légères sur les nageoires.
Le poisson possède deux lignes latérales, une ligne supérieure et une ligne inférieure, avec les quatre à sept barres mentionnées plus tôt centrées entre elles.
L’apparence du poisson peut changer en quelques secondes lors de comportements agressifs, compétitifs ou de reproduction.
La couleur de base du corps est généralement gris-jaunâtre à brun.
Pendant la période de frai, le côté ventral de la femelle devient sombre et le corps prend une couleur blanchâtre.
Les mâles matures dominants sont généralement foncés avec une grande bosse occipitale.
Il peut y avoir plusieurs bandes transversales à faible contraste sur les côtés du corps, mais elles sont pour la plupart pratiquement invisibles.
Des taches bleuâtres ou jaunâtres sont dispersées dans tout le corps et à la base de la queue se trouve une tache noire à fort contraste.
En aquarium, les mâles deviennent rarement complètement noirs et cela tient probablement au fait que la domination en aquarium est plus facile à atteindre et à maintenir.
Un mâle dominant obtient ainsi une sorte de monopole dans un aquarium qui doit être moins défendu.
Dans la nature, en revanche, de nouveaux rivaux rôdent constamment.
C’est pourquoi les animaux là-bas conservent les couleurs d’avertissement foncées.
–
SIGNES DISTINCTIFS
Pour identifier le genre Herichthys, il faut repérer la présence des six à sept barres verticales sur la face centrale postérieure du poisson.
Pendant la saison de reproduction, la coloration des poissons change pour se préparer à la reproduction, présentant une dichromatisme sexuel avec ce nouveau look de reproduction.
Herichthys minckleyi se distingue des autres espèces du groupe par sa tête plus longue, son museau plus long et ses nageoires dorsale et anale plus courtes.
Elle peut être identifiée à seize épines et onze rayons sur la nageoire dorsale, avec cinq épines sur la nageoire anale.
Pour identifier le genre Herichthys, on peut observer les six à sept barres verticales présentes sur la face postérieure médiane du poisson.
Pendant la saison de reproduction, la coloration du poisson change en prévision de la reproduction, présentant un dichromatisme sexuel avec cette nouvelle apparence nuptiale.
Herichthys minckleyi se distingue des autres espèces du groupe par sa tête plus longue, son museau plus long et ses nageoires dorsale et anale plus courtes.
Il est reconnaissable à ses seize épines et onze rayons sur la nageoire dorsale, et à ses cinq épines sur la nageoire anale.
–
Caractéristiques clés de Herichthys minckleyi
- La carrure est haute, ovale et aplatie latéralement ;
- La coloration de base de son corps est généralement jaunâtre gris à brunâtre ;
- Pendant les soins de la couvée, le ventre des femelles s’assombrit et le corps s’encadre de façon claire (blanchâtre) ;
- Les mâles dominants sexuellement matures sont généralement de couleur foncée ;
- sur les côtés du corps, il peut y avoir plusieurs bandes transversales (faibles), souvent invisibles ;
- Au centre des flancs du corps passe une bande longitudinale noire, dissoute en plusieurs endroits de tailles différentes ;
- À la base de la nageoire caudale se trouve une tache noire contrastante ;
- la tête est relativement grande, la terminale de la bouche et les lèvres sont épaissies.
–
DIFFERENCIATION
Pour identifier le genre Herichthys, on peut observer les six à sept barres verticales présentes sur la face postérieure médiane du poisson.
Pendant la saison de reproduction, la coloration du poisson change en prévision de la reproduction, présentant un dichromatisme sexuel avec cette nouvelle apparence nuptiale.
Herichthys minckleyi se distingue des autres espèces du groupe par sa tête plus longue, son museau plus long et ses nageoires dorsale et anale plus courtes.
Il est reconnaissable par :
- Ses seize épines et onze rayons sur la nageoire dorsale ;
- Ses cinq épines sur la nageoire anale.
Ce cichlidé se distingue des autres membres du genre par son polymorphisme, avec la présence, en sympatrie, d’individus présentant deux morphologies distinctes (l’une allongée et élancée, l’autre beaucoup plus haute).
Les trois types de Herichthys minckleyi individus possèdent les caractéristiques suivantes :
- Deux types de dents pharyngiennes : molariformes et papillaires ;
- Les dents orales sont coniques.
- Nageoire dorsale : XVI rayons, 10-11 branches ;
- Nageoire anale : V rayons, 8-9 branches ;
- 14 rayons pectoraux ;
- 11-12 arcs branchiaux ;
- 29 vertèbres ;
- Le pédoncule caudal est d’une longueur égale ou supérieure à sa hauteur ;
Les adultes présentent un dichromatisme sexuel lors de la reproduction et des soins parentaux :
Le mâle est vert foncé à noir avec des extrémités bleues sur le corps et les nageoires ;
La femelle est blanche avec des marques noires sur les flancs.
Chez les individus sexuellement inactifs, la coloration est très variable et ils peuvent changer de couleur très rapidement en fonction de leur humeur, de leur environnement social, etc.
La couleur de base du corps varie du gris clair à l’ocre, en passant par le jaune verdâtre jusqu’au vert foncé ou au gris.
Ils présentent des marques noires sur le corps, bien que celles-ci varient considérablement d’un individu à l’autre.
La plupart ont des taches bleues ou jaunâtres sur les flancs, la tête et les nageoires.
Dans certaines localités de la vallée de Cuatro Ciénegas, Herichthys cyanoguttatus est très semblable à Herichthys minckleyi et vit en sympatrie avec cette dernière.
Parmi les différences caractéristiques entre les deux espèces, on note :
- la forme des dents orales (bicuspides chez Herichthys cyanoguttatus et coniques chez Herichthys minckleyi) ;
- la forte présence de dents pharyngiennes molariformes chez Herichthys cyanoguttatus, alors que seule une des formes de Herichthys minckleyi en possède ;
- la coloration corporelle des individus adultes en période de reproduction (les femelles et les mâles de Herichthys cyanoguttatus présentent une partie antérieure blanche et une partie postérieure foncée, tandis que les femelles de Herichthys minckleyi sont blanches à l’exception de quelques taches noires sur les flancs et à la base de la nageoire caudale ; les mâles sont noirs ou très foncés).
–
DUREE DE VIE
L’espérance de vie de ces poissons varie de 6 → 8 ans.
–
DIMORPHISME SEXUEL
Déterminer les mâles des femelles n’est pas chose simple, surtout tant que les poissons n’ont pas atteint leur taille adulte.
Cependant, chez tous les Herichthys, il y a un certain dimorphisme sexuel.
En plus d’être légèrement plus gros que la femelle, les Herichthys mâles ont souvent des nageoires dorsale et anale visiblement plus longues et plus pointues, et les mâles plus âgés peuvent développer une bosse nucale en vieillissant.
Les Herichthys minckleyi ont des particularités propres :
- Lors de la reproduction, le mâle Herichthys minckleyi devient noir tandis que la femelle Herichthys minckleyi devient blanche avec des taches noires.
- Outre les différences de couleur, il existe également une différence de taille : Les mâles sont en moyenne 10 % plus grands et présentent souvent une bosse frontale.
- Les mâles dominants se distinguent normalement par la présence de cette bosse nucale sur le front mais aussi par une coloration plus foncée.

Couple de Herichthys minckleyi : Mâle à gauche & Femelle à droite.
Les mâles adultes passent du vert foncé au noir complet, avec des taches bleu clair réparties sur la tête, les flancs et les nageoires verticales.
En dernier ressort, il reste deux solutions :
- Observer quand un couple s’est formé ;
- Généralement le mâle est plus grand que la femelle ;
- La méthode de la « Retournette » ou « Venting » qui permet d’observer les orifice génitaux des poissons. Cette méthode demande un peu d’expérience et de pratique pour l’éleveur, ne serait-ce que, déjà, pour attraper ces poissons qui dans ces conditions se montrent vifs pour s’échapper !
La retournette, doit être pratiquée après la maturation sexuelle (TL = 13 centimètres), afin d’être plus fiable.
Cette méthode est certainement la meilleure à condition de savoir bien observer : l’organe excréteur et reproducteur de chaque poisson.
Þ Chez le mâle, cet organe est en forme de « oo »,
Þ Chez la femelle, cet organe est en forme de« o0 ».
C’est la pratique la plus sûre de différenciation entre les sexes, et
–
COMPORTEMENT
Les Herichthys minckleyi mènent un mode de vie sociable.
Herichthys minckleyi est un poisson grégaire, formant des groupes et des colonies pour se nourrir et vivre dans les sources.
Les territoires hors période de reproduction sont occupés par un grand nombre de poissons, la plupart étant ignorés par le mâle dominant.
Ce dernier se distingue généralement par la présence d’une bosse proéminente sur son front, ainsi que par une coloration plus foncée.
On trouve une grande quantité de poissons dans ces territoires, principalement des femelles ou des mâles non dominants.
Dans l’aquarium, les poissons peuvent être gardés en groupes, où il devrait y avoir 3 à 4 femelles pour un mâle.
Dans le cas de la présence de plusieurs mâles ensemble, l’aquarium doit être suffisamment spacieux et comporter de nombreux abris pour réduire le risque de conflits entre eux.
Dans les territoires, on trouve une grande quantité de poissons.
La majorité d’entre eux seront des cichlidés femelles ou des mâles non dominants.
Un mâle dominant se distingue normalement par la présence d’une bosse nucale le front, ainsi que d’une coloration plus foncée.
Les territoires des poissons non reproducteurs seront habités par une grande quantité de poissons, dont la plupart seront ignorés par le mâle dominant qui s’y trouve.
D’autres cichlidés du Mexique et/ou d’Amérique centrale peuvent être tolérés, mais lors de la reproduction ces poissons il sera préférable que les Herichthys minckleyi soient gardés seuls car, dans ces moments, ce sont de vrais terreurs qui demandent un aquarium strictement spécifique.
–
CARACTERE
Maintenir cette espèce de cichlidé en captivité et en obtenir la reproduction et la croissance de sa descendance n’est pas chose facile , voire plutôt difficile !
Herichthys minckleyi est généralement un poisson territorial, surtout pendant la saison de frai.
Il s’agit d’une espèce très sociale dotée de systèmes hiérarchiques et de dominance complexes.
En aquarium, la reproduction nécessite obligatoirement la mise à disposition d’un espace suffisant et des zones où la femelle et les autres individus peuvent échapper en cas d’agressivité des mâles.
L’agressivité entre mâles est fréquente, entraînant souvent la mort des mâles non dominants, voire de certaines femelles.
Herichthys minckleyi est parfois discret et se cache volontiers dans le décor pour échapper aux combats qui existent entre les spécimens.
D’autres cichlidés du Mexique et d’Amérique centrale peuvent être tolérés avec les Herichthys minckleyi, mais lors de leur reproduction, ces poissons doivent être gardés seuls.
Ils manifestent souvent un comportement agressif envers leurs congénères et autres poissons, en particulier s’ils sont hébergés dans un aquarium trop petit pour leurs évolutions : Herichthys minckleyi aime les grands espaces !

Souvent, en captivité, ils deviendront de vrais terreurs ce qui imposera de leur donner un aquarium spécifique pour éviter les conflits et dans ces conditions, un aquarium spécifique avec suffisamment d’espace et de cachettes est recommandé.
Herichthys minckleyi est dans une certaine mesure un poisson grégaire, formant des groupes et des colonies pour se nourrir et vivre dans les sources.
Les zone franches dans lesquels des territoires de reproducteurs ne sont pas installés, seront habités par une grande quantité d’espèces de poissons, dont la plupart seront ignorés par les Herichthys minckleyi.
Dans les territoires, on trouve une grande quantité de poisson.
La majorité d’entre eux seront des cichlidés femelles ou des mâles non dominants.
Même un mâle dominant, noir foncé, qui s’y trouverait, ne manifestera aucune agressivité.
Dans un aquarium, contrairement au milieu naturel qui offre plus de possibilités de fuites et d’échappatoires, les mâles rivaux sont éliminés prématurément, et les mâles subordonnés ont du mal à affirmer leur statut social et s’imposer.
En aquarium, les mâles deviennent rarement complètement noirs et cela tient probablement au fait que la domination en aquarium est plus facile à atteindre et à maintenir car les rivaux sont éliminés prématurément, et les mâles subordonnés ont du mal à progresser en statut social.
Un mâle dominant obtient ainsi une sorte de monopole dans un aquarium qui est un espace restreint, moins peuplée et qui a par conséquent moins besoin d’être défendu.
Dans leur milieu naturel en revanche, le plus souvent de jeunes mâles Herichthys minckleyi rivaux rôdent constamment et les mâles dominants sont sur leur garde ce qui explique que ces animaux conservent les couleurs d’avertissement foncées.
D’après les éleveurs qui ont fait l’expérience de la maintenance de Herichthys minckleyi en aquarium avec les meilleurs conditions possibles, il a pu être observé également une grande différence dans le comportement reproductif du mâle Herichthys minckleyi en aquarium : Il semblerait qu’il se montre bien plus attentionné que dans la nature.
Enfin, comparés au cichlidé monogame du Rio Grande, Herichthys cyanoguttatus, les mâles Herichthys minckleyi sont moins présents dans leurs nids et affichent des taux de comportement de défense et d’agressivités plus faibles, et les femelles compensaient l’absence de leurs partenaires en assurant un niveau de défense plus élevé sur leur descendance.
–
COHABITATION
Herichthys minckleyi est dans une certaine mesure un poisson sociable, formant des groupes et des colonies pour se nourrir et vivre dans les sources.
Les territoires non reproducteurs sont habités par une grande quantité de poissons, dont la plupart seront ignorés par le mâle dominant.

–
LES POISSONS DE QUATRO-CIENNEGAS
CYPRINIDAE – Cyprinidés
Cyprinella xanthicara (MINCKLEY & LYTLE) – Méné Cuatrociénegas / Sardinita de Cuatrociénegas |A| |P| Endémique.

Dionda épiscopa (GIRARD) – roundnose minnow / Sardinita nez rond |M| |P| Natif

Cyprinus carpio (LINNAEUS) – common carp / Carpe commune. Collectée sur un site dans les années 1990, mais jamais après. Éteinte ? Non-native

–
CHARACIDAE – Characins / Sardines
Astyanax mexicanus (FILIPPI) – Tetra mexicain / sardina de Mexico |P| Natif

–
ICTALURIDAE – Bullhead Catfishes / Bagres
Ictalurus lupus (GIRARD) – poisson-chat d’amont / lobo du sac |P| Natif

Pylodictis olivaris (RAFINESQUE) – flathead catfish /Bagre à tête plate |P| Natif

–
POECILIDAE – Livcbearers / Viviparos
Gambusie longispinis (MINCKLEY) – Cuatrociénegas gambusia / Guayacón de Cuatrociénegas Endémique

Gambusia marshi (MINCKLEY & CRADDOCK) – robust gambusia / Goyacón des nageurs |P| Natif


Xiphophorus gordoni (MILLER & MINCKLEY) – Cuatrociénegas platyfish / Espada (o Platy) de Cuatrociénegas |A| |P| Endémique

–
FUNDULIDAE – Topminnows
Lucania interioris (HUBBS & MILLER) – Cuatrociénegas killifish / Sardinilla de Cuatrociénegas |A| Endémique

–
CYPRINODONTIDAE – Pupfishes
Cyprinodon atrorus (MILLER) – bandfin pupfish / chiot de Sacquet |A| |P| Endémique

Cyprinodon bifasciatus (MILLER) – chioton / Cachorrito de Cuatrociénegas |A| |P| Endémique

–
CENTRARCHIDAE – Crapets / Mojarras et Lobinas
Lepomis megalotis ssp. – Cuatrociénegas sunfish / Mojarra de sol de Cuatrociénegas |P| Endémique – non décrit

Micropterus salmoides ssp. – Cuatrociénegas bass / Lobina o Robalo de Cuatrociénegas |P| Endémique – Non décrit

Micropterus floridianus (LESUEUR 1822) – Bass de Floride / Robalo de Floride. Marqueurs génétiques détectés avec des indications d’introgression aux populations du lobine/vol indigène. Non natif

–
PERCIDAE – Perches and Darters / Perches
Etheostoma lugoi (NORRIS & MINCKLEY) – Quatro ciennegas darter / Fourchettes |A| |P| Endémique

Etheostoma segrex (NORRIS & MINCKLEY) – Rivière Salée darter / Dard del Salado |A| Endemique de la partie haute de la rivière Salée des Nageurs peu en dehors de la vallée des Quatro cienégas)

–
CICHLIDAE – Cichlidés / Cíclidos
Herichthys cyanoguttatus (BAIRD & GIRARD) – Cichlidé du Rio Grande / Mojarra del Norte |P| (indigène ou peut-être un des premiers envahisseurs via les premiers canaux qui extrayaient l’eau du bassin des Quatro ciénegas anciennement fermé / native ou peut-être une envahissante il y a de nombreuses années via les premiers canaux qu’ils ont extrait l’eau de l’avant fermée vallée des Quatro cienégas)

Herichthys minckleyi (KORNFIELD & TAYLOR) – Cuatro ciénegas cichlid / Mojarra de Cuatro ciénegas |P| Endémique
Hemichromis guttatus (GÜNTHER) – Jewel cichlid / Poisson bijou (non-natif) => Cichlidé africain

Oreochromis aureus (STEINDACHNER) – Blue Tilapia / Tilapia bleu (non-natif) => Cichlidé africain

Au total, 21 espèces de poissons peuplent les eaux de Cuatro Ciénégas !
–
EAU
VALEURS D’EAU MESUREES DANS LA NATURE
L’environnement où vit Herichthys minckleyi est très particulier car il est unique.
Les sources de Cuatro Cienegas se situent au cœur d’un paysage désertique foisonnant de plantes vasculaires et de cactus nains, et sont entourées d’impressionnantes crêtes rocheuses dénudées.
On constate que les précipitations annuelles dans cette zone n’excèdent pas 300 millimètres par an.
L’eau de pluie est rapidement absorbée par le sol calcaire poreux de la vallée.
Il est constitué de sources d’eau chaude et cristalline, dont la visibilité dépasse généralement vingt mètres et dont la température est très stable.
Des mesures effectuées sur une période de dix ans dans la Laguna del Mojarral ont montré une température comprise entre 33,5°C et 34,0°C, soit une variation de seulement 0,5°C (ARNOLD, 1972), avec des variations maximales de 5°C en surface.
La profondeur maximale de ces sources est actuellement d’environ cinq mètres (Laguna del Mojarral, Poza de la Becerra), et leur surface est relativement réduite.
La poza El Mojarral mesure environ 30 x 70 mètres, tandis que la poza La Becerra mesure environ 50 x 100 mètres et la Poza Churince environ 15 x 30 mètres : ce sont des espaces assez petits et très contraints.
Ces poissons préfèrent l’eau thermale de ces pozas (bassins ou piscines naturels) où l’on trouve de l’eau cristalline dont la visibilité est élevée, souvent supérieure à 20 mètres et avec une température comprise entre 30 et 34°C.
Dans ces pozas, le pH est légèrement alcalin.
Les mesures effectuées à plusieurs reprises par ARTIGUAS AZAS ont varié de 7,6 à 7,8, avec une dureté moyenne d’environ 55 degrés allemands.
On retiendra les éléments suivants pour une maintenance en aquarium :
- Températures minimale à maximale : 12 à 35°C ;
- PH : 7,5 – 8,5 ;
- DH :
Dans l’aquarium de ces poissons, il faudra maintenir des températures comprises entre 25 et 30°C et une eau alcaline avec une dureté moyenne à élevée.
Ces niveaux d’eau sont très stables (variation annuelle inférieure à un degré centigrade), ainsi que le niveau et la qualité de l’eau sans grandes variations (WINSBOROUGH 1990 ; HENDRICKSON & Al., 2005).
L’eau douce doit impérativement être évitée car elle rend ces animaux sensibles aux maladies : Herichthys minckleyi apprécie une eau à pH neutre ou légèrement basique et un peu dure.
La température de l’eau varie fortement entre les bassins et les sources chaudes et froides émergent souvent à quelques mètres les unes des autres, se fondant en ruisseaux, rivières et marais.
–
ENVIRONNEMENT & ZONE DE VIE
Herichthys minckleyi vit dans des environnements qui ressemblent à des les lagons, alimentés par des sources d’eau pures, fraîches à chaudes, dans de petits étangs et ruisseaux aux eaux très claires pouvant être très légèrement salées avec des profondeurs allant jusqu’à 7 mètres.
Herichthys minckleyi habite toutes les couches d’eau et a surtout besoin de grands espaces pour évoluer.
ALIMENTATION
EN MILIEU NATUREL
Le régime alimentaire des Herichthys minckleyi se compose de plantes, d’algues, de détritus, de mollusques (escargots de la famille des Hydrobiidae), d’autres invertébrés aquatiques et terrestres, et de poissons.
Cependant, il faut savoir que seuls les individus de forme molariforme peuvent consommer des escargots, mais en ce qui concerne les autres, les régimes alimentaires se chevauchent considérablement (SMITH 1982 ; HULSEY & Al. 2005).
Cependant, des études sur les isotopes stables du carbone et de l’azote ont révélé des différences de régime alimentaire entre les différents morphotypes et entre les adultes et les juvéniles.
La spécialisation alimentaire du cichlidé Herichthys minckleyi, présentant un polymorphisme trophique, a été étudiée à partir de son contenu stomacal.
Les Herichthys minckleyi ont été classés selon la morphologie de leurs mâchoires pharyngiennes :
- Molariformes: Les mâchoires pharyngiennes de type « molariforme » présentent des dents aplaties et élargies, tandis que celles de type « papilliforme » se caractérisent par la présence de petites dents en forme de crayon sur la mâchoire pharyngienne (LIEM & KAUFMAN, 1984) ;
- Papilliformes;
- Piscivores et Indéterminées.
La présence de dents pharyngiennes aplaties et élargies a permis de classer Herichthys minckleyi comme molariformes, tandis que la présence de petites dents pharyngiennes fines comme un crayon a permis de le classer comme papilliformes.

Différence entre type mollariforme & Papiliforme.
Les individus indéterminés (longueur standard < 50 millimètres ) n’ont pas été rattachés à l’un des deux morphotypes les plus grands.
Cependant, aucune différence significative de régime alimentaire n’a été mise en évidence.
Des études antérieures indiquent que les détritus, les algues, les macro-invertébrés (SAGE & SELANDER, 1975 ; SMITH, 1982) et les coquilles d’escargots (SAGE & SELANDER, 1975 ; SMITH, 1982) sont consommés aussi bien par les molariformes que par les papilliformes.
Malgré ce chevauchement qualitatif de leur régime alimentaire, il a été suggéré que la mâchoire pharyngienne des papilliformes favorise le broyage des arthropodes et des végétaux.
On suppose que les dents massives des molariformes sont adaptées au broyage des coquilles d’escargots résistantes (KORNFIELD & KOEHN, 1975 ; SAGE & SELANDER, 1975 ; HULSEY & Al., 2005).
Une quantification plus poussée de l’utilisation des proies permettrait de mettre en évidence des différences de niche morphologique.
Les différents morphotypes pharyngiens différent par leur contenu intestinal.
La contribution volumétrique totale, en pourcentage, des différents types de proies, par ordre décroissant d’importance, dans l’intestin de Herichthys minckleyi était la suivante :
- Débris végétaux : 33,6 %,
- Débris : 30 % ;
- Travertin : 14,4 % ;
- Coquilles d’escargots : 11,6 % ;
- Odonates : 20 % ;
- Poissons : 14 % ;
- Palaemonetes sp. : 12 % ;
- Chironomidés : 12 % ;
- Hyallela sp. : 11 % ;
- Ostracodes : 10 % ;
- Hyménoptères : 7 % ;
- Coléoptères : 4 % ;
- Ephémères : 2 % ;
- Lépidoptères : 2 % ;
- Ecailles de poissons : 2 % ;
- Acariens : 1 % ;
- Diptères autres que les Chironomidés : 1 % ;
- Hémiptères : 1 %.
Les arthropodes représentent une faible part du volume intestinal total.
Les Chironomidae, les Hyallela et les Ostracoda étaient les arthropodes les plus fréquemment ingérés, mais leur contribution au contenu intestinal de tout type de mâchoire pharyngienne n’était pas substantielle.
Les arthropodes ne sont pas consommés plus fréquemment par les Herichthys minckleyi molariformes, ni par les papilliformes, bien que les molariformes aient eu tendance à ingérer ce type de proie plus fréquemment que les papilliformes.
De plus, les poissons ne représentent qu’une faible part du régime alimentaire de tous les morphotypes.
La grande majorité du volume intestinal est composée de coquilles d’escargots, de travertin, de détritus et de débris végétaux.
L’ingestion d’escargots est répartie de manière non aléatoire parmi les morphotypes de mâchoires.
Les coquilles d’escargots représentent un faible pourcentage du contenu intestinal des papilliformes (< 0,5 %) et seuls trois intestins de papilliformes contenaient des opercules.
Chez les molariformes, 28 % du contenu intestinal total est constitué d’escargots.
Les papilliformes consomment davantage de matière végétale que les molariformes.
Leur capacité à broyer et manipuler efficacement la matière végétale, comparativement aux molariformes (HULSEY & Al., 2005), pourrait s’expliquer par la morphologie plus fine de leur mâchoire pharyngienne et leurs petites dents pointues.
Un broyage plus complet des détritus végétaux par les papilliformes permettrait une extraction plus efficace des nutriments contenus dans ces aliments difficiles à digérer (HORN, 1989).
L’utilisation des proies différait considérablement selon les morphotypes pharyngiens d’Herichthys minckleyi.
Les plantes et les escargots n’étaient pas consommés de manière intensive par les Herichthys minckleyi de taille indéterminée.
Le même pourcentage de matière végétale retrouvé dans les intestins des molariformes était également présent dans ceux des individus de taille indéterminée, et ces derniers ne broyaient jamais d’escargots.
Les mâchoires pharyngiennes des Herichthys minckleyi de moins de 50 millimètres de longueur totale ne sont pas encore soudées et leurs dents et muscles ne sont pas complètement développés (STEPHENS & HENDRICKSON, 2001).
En raison de ces caractéristiques, le broyage des plantes et l’écrasement des escargots ne constituent probablement pas des moyens efficaces de traitement des proies pour les Herichthys minckleyi de taille indéterminée.
Il a été démontré expérimentalement que les deux formes d’Herichthys minckleyi, molariforme et papilliforme, privilégient la consommation d’arthropodes plutôt que le broyage d’escargots (LIEM & KAUFMAN, 1984).
L’exploitation intensive des arthropodes par Herichthys minckleyi dans son milieu naturel n’est pas fortement suggérée par la proportion volumétrique ni la fréquence de ce type de proie dans son régime alimentaire.
Cuatro Ciénegas se caractérise par une très faible abondance d’arthropodes aquatiques (DINGER, 2001), et la faible prévalence de ces proies dans le régime alimentaire d’Herichthys minckleyi pourrait être une conséquence de cette faible disponibilité.
Les spécialisations mentionnées sont principalement exprimées en période de pénurie alimentaire.
Cependant, les animaux ont conservé leur préférence pour les insectes aquatiques et les larves.
Cela devient évident lorsqu’ils sont gardés en aquarium et qu’on leur propose un menu varié.
Dès que les mangeurs d’escargots peuvent avoir quelque chose de mieux, c’est-à-dire quelque chose qui demande moins d’énergie à atteindre et ou a une valeur nutritionnelle plus élevée, ils ignorent rapidement les escargots.
La proportion d’escargots dans le menu d’un molluscivore varie, saisonnière ou autrement, selon la disponibilité alimentaire.
Les animaux utilisent principalement leur spécialité en période de rareté, mais dans les zones désertiques, c’est bien sûr souvent la plus grande partie de l’année.
L’avantage de se nourrir de plantes ou d’escargots pourrait être accentué par la faible densité d’arthropodes à Cuatro Ciénegas.
Les papilliformes consommaient davantage de matière végétale que les molariformes.
Leur capacité à broyer et manipuler efficacement la matière végétale, comparativement aux molariformes (HULSEY & Al., 2005), pourrait s’expliquer par la morphologie plus fine de leur pharyngien et leurs petites dents pointues.
Un broyage plus complet des détritus végétaux par les papilliformes permettrait une extraction plus efficace des nutriments contenus dans ces aliments difficiles à digérer (HORN, 1989).
Une différenciation plus fine des niches trophiques au sein d’Herichthys minckleyi pourrait être mise en évidence par une catégorisation taxonomique plus précise de la matière végétale ingérée par les deux morphotypes.
Les escargots étaient rarement, voire jamais, broyés par les papilliformes, mais les trois espèces d’escargots robustes et communes de Cuatro Cienegas sont fréquemment broyées par les molariformes.
Mexipyrgus churinceanus est l’espèce la plus fréquemment ingérée, mais Mexipyrgus churinceanus et Mexipyrgus minckleyi étaient également souvent broyés par les molariformes.
Aucune corrélation n’a été observée entre la taille de la coquille des molariformes et le nombre d’escargots broyés.
De plus, moins de la moitié des molariformes examinés broyaient des escargots, et seulement 28 % du contenu intestinal des molariformes était constitué de coquilles d’escargots.
Cette faible consommation d’escargots pourrait s’expliquer par la robustesse de leur coquille (HULSEY & Al., 2005), ce qui inciterait les molariformes à consommer d’autres proies lorsque celles-ci sont disponibles.
La disponibilité des proies tout au long de l’année à Cuatro Cienegas devrait être évaluée plus en détail car les comportements alimentaires des morphotypes Herichthys minckleyi varient de façon saisonnière (COHEN & Al., 2005).
Étant donné que la fréquence des morphotypes varie selon les habitats (HENDRICKSON & Al., sous presse), un examen plus approfondi des différences de disponibilité des proies entre les bassins pourrait également fournir des informations sur la spécialisation trophique de Herichthys minckleyi.
–
EN AQUARIUM
Bien que les habitudes alimentaires des poissons soient fortement corrélées à la morphologie des trois formes différentes, les analyses ont montré que les spécimens sauvages avaient invariablement qu’un seul type de des aliments dans leur tube digestif et sous conditions d’abondance alimentaire.
Þ Toutes sortes d’aliments sont consommés…ce qui facile grandement l’éleveur de ces poissons en aquarium !
Les études ont montré que les spécialisations mentionnées s’expriment principalement lors de la pénurie alimentaire et que ces animaux ont conservé leur préférence pour les insectes aquatiques et les larves.
Ce constat devient parfaitement évident lors de la maintenance des Herichthys minckleyi en aquarium et lorsque des menus variés leur sont proposés, quels que soient leurs types, ces poissons ne font pas les difficiles.
Dès que les mangeurs d’escargots peuvent obtenir quelque chose de mieux, à savoir quelque chose qui prend moins d’énergie à saisir et ou une valeur nutritive plus élevée, ils ignorent rapidement les escargots.
La proportion d’escargots dans le menu d’un molluscivore varie, selon la saison ou selon la disponibilité de la nourriture.
Les animaux utilisent donc leur spécialisation surtout en période de pénurie, mais dans les zones désertiques, c’est bien sûr généralement la plus grande partie de l’année.
Les caractéristiques qui séparent ces trois types ont une composante héréditaire et ne sont pas seulement le résultat de la plasticité phénotypique (variation due à des facteurs environnementaux), mais ces caractéristiques sont déclenchées par des facteurs environnementaux.
Par exemple, un molluscivore est plus susceptible de développer des dents en forme de molaire lorsqu’il est confronté à des aliments durs.
À l’inverse, cependant, il n’est pas vrai que les dents en forme de molaire disparaissent également une fois que les animaux ne reçoivent que de la nourriture molle.
Un autre fait qui montre la composante héréditaire est qu’un détracteur ne développera jamais de dents en forme de molaire, même si nous les mettons au régime des escargots.
Le poisson n’a tout simplement pas les bons gènes pour développer des dents en forme de molaire.
–
REGIME
Les dents molariformes permettent de broyer les escargots, tandis que peu de poissons papilliformes en consomment à l’état sauvage.
Inversement, les poissons papilliformes broient finement les plantes pour se nourrir, ce dont les poissons molariformes sont incapables.
Fait remarquable, la force musculaire nécessaire au broyage des escargots chez le poisson molariforme Herichthys minckleyi est considérable, même comparée à celle d’autres poissons mollusques-nourrisseurs.
Ceci pourrait être dû à des interactions prédateur-proie co-évolutives entre les escargots résistants de la région de Cuatro Ciénegas et les puissants muscles pharyngiens de Herichthys minckleyi.
Ces deux morphotypes se nourrissent dans des micro-habitats différents et présentent également des comportements alimentaires distincts.
Cette distinction réduit probablement la compétition pour les ressources au sein de l’espèce.
Ce polymorphisme persiste au sein de la population, au lieu de se confondre, en raison de cette compétition réduite entre les différents morphotypes, et est considéré comme une première étape importante vers la spéciation.
Les cichlidés en général sont connus pour leur spéciation rapide, d’où l’intérêt porté à ces formes divergentes tout en conservant une homogénéité génétique.
En aquarium, ils peuvent manger des flocons et d’autres aliments préparés ou vivants.
Autrement dit, ils mangent tout ce qu’ils trouvent.
On a également constaté en laboratoire qu’en période d’abondance alimentaire, la forme molluscivore considère les escargots comme la nourriture la moins désirable.
Ceci explique la présence d’arthropodes dans l’intestin des trois morphotypes. Cela montre aussi que lorsqu’une occasion se présente, ils ne la laissent pas passer.
C’est seulement lorsque la nourriture se fait rare, ce qui est le cas la plupart du temps étant donné que les sources de Cuatro Cienegas sont presque des déserts sous-marins, que les morphotypes se concentrent sur leur mode d’alimentation particulier.
–
AQUARIUM
Herichthys minckleyi est un poisson benthopélagique du Mexique, endémique du bassin endoréique de Cuatro Ciénegas à Coahuila (versant Atlantique mexicain) et, en plus d’être en voie de disparition, cette espèce rare est difficile à élever en aquarium.
Herichthys minckleyi est un cichlidé très agressif, selon certains, ce serait même le cichlidé méso-american le plus agressif qui puisse exister !
Conserver ce poisson en captivité dans un aquarium ne sera possible que dans les plus grands aquariums, c’est à dire dans un bac mesurant trois mètres et plus.
L’élevage durable en captivité n’est possible que dans les plus grands aquariums, c’est-à-dire dans des bacs mesurant au moins trois mètres et plus.
Les types 1 et type 2 sont mieux tenus en groupe et de préférence avec différentes catégories d’âge mélangées.
Pour tous les types d’Herichthys minckleyi, il faudra prévoir un aménagement spécifique du bac avec la mise ne place de cachettes suffisantes et, surtout, adaptées qui pourront servir de refuges pour les poissons de rangs inférieurs, mais aussi pour les opportunités de reproduction nécessaires.
Après tout, nous avons affaire à un pondeur/éleveur en grotte.

Les Herichthys minckleyi de type 3 sembleraient mieux vivre en binôme et avec d’autres grands cichlidés et souvent, il s’avère que ce type d’Herichthys minckleyi semble apprécier de mener un style de vie plus solitaire.
Autre particularité, il faudra prendre en compte le type de Herichthys minckleyi et, en ce qui concernent les type 1 et type 2, il faudra privilégier une maintenance en groupe combinant, de préférence, différentes catégories d’âge ensemble.
–
CONFIGURATION DE L’AQUARIUM
Compte tenu de la nature exceptionnel de l’environnement d’origine de ces poissons endémiques, le défi pour l’éleveur sera d’offrir le maximum de critère que les eaux de Cuatro Cienegas offrent.
–
DECOR & AQUARIUM BIOTOPE
Le choix d’un très grand bac est la condition de base pour le succès de la maintenance des Herichthys minckleyi.
Pour bien faire, il serait idéal de reconstruire au maximum le biotope d’origine de ces poissons mais un très grand bac avec un décor rocheux assez épuré tendra vers l’idéal et reproduira assez facilement le milieu aquatique d’origine de ces cichlidés.
Il y pour l’éleveur un véritable défi à réaliser pour ce bac destiné aux Herichthys minckleyi et dont le biotope est aussi particulier coque le sont les qualités de l’eau.
–
PLANTES
La présence de plantes dans le bac de ces poissons n’est pas une obligation puisque dans leur milieu naturel, les plantes sont peu nombreuses.

–
ECLAIRAGE
Un éclairage normal suffira pour mettre en valeur ces poissons.
–
FILTRATION
Une filtration de haute qualité de l’eau et une aération améliorée sont nécessaires, ainsi qu’un changement hebdomadaire d’un tiers de sa partie par de la fraîche.
Les Herichthys minckleyi sont relativement de gros poissons qui consomment donc une grande quantité de nourriture et produisent une grande quantité de déchets qui devront être éliminés en conséquence avec l’obligation d’avoir en permanence une eau toujours la plus pure comme c’est le cas dans la nature pour ce poisson.
Par conséquent, si un effort particulier doit être porté, c’est sur la qualité de l’eau et donc la filtration !
Cette filtration doit être suffisamment puissante pour traiter les produits azotés résultants.
La filtration du doit être robuste pour ces poissons et l’équipement de filtration du bac devra pouvoir effectuer environ 6 à 10 fois le renouvellement du réservoir par heure quelle que soit la taille du bac de ces Herichthys.
Même avec un faible taux d’occupation, une forte filtration, les changements d’eau fréquents et réguliers.
Plus que tous les autres Herichthys, les Herichthys minckleyi sont sensibles aux accumulations de nitrates et de grands changements d’eau seront donc également nécessaires, de l’ordre de 30% hebdomadaire et avec une eau qui aura été préparée, c’est-à-dire déchlorée, oxygénée…
Si ces exigences de soins ne sont pas respectées, les Herichthys minckleyi courent le risque de dépérir progressivement avec souvent une issue mortelle pour ces pensionnaires exceptionnels.

–
REPRODUCTION
Herichthys minckleyi est réputé pour former des couples stables et prodiguer des soins biparentaux à sa progéniture
Herichthys minckleyi atteint la maturité sexuelle à l’âge d’un an : La taille de maturité est d’environ 6 centimètres de longueur museau-cloaque (LMC) chez les femelles.
Cette taille de reproduction sera atteinte avant la deuxième année d’âge de la femelle.
Les Herichthys minckleyi sont polygames.
–
Taux de croissance
La croissance est influencée par l’alimentation et la température.
Des données préliminaires sur l’âge et la croissance sont disponibles grâce à des études non publiées de HENDRICKSON (2005).
En laboratoire, la croissance est très variable entre les individus d’une même génération.
Certains individus ont été recapturés six ans après avoir été marqués dans différents habitats de Cuatro Ciénegas.
Un spécimen en laboratoire est âgé d’au moins 11 ans (HENDRICKSON, données non publiées).
Le taux de croissance d’un échantillon de plus de 200 individus recapturés à Cuatro Ciénegas (plus de trois fois sur plusieurs années) était en moyenne de 0,02 millimètres par jour.
–
REPRODUCTION ET SOINS PARENTAUX
Les Herichthys minckleyi sont la seule espèce de leur genre à présenter un dichromatisme sexuel, c’est-à-dire que les mâles et les femelles présentent des motifs de coloration nettement différents lors de l’accouplement pour la reproduction.
Cette caractéristique unique a évolué après la divergence de l’espèce par rapport aux autres Herichthys vivants.
Les femelles deviennent généralement blanches avec des marques noires sur les flancs, tandis que les mâles arborent une coloration vert foncé ou noire unie avec des taches bleuâtres pâles sur les nageoires.

Cependant, d’autres variations de couleurs existent, et les individus n’expriment pas toujours complètement la coloration nuptiale : De nombreux changements de coloration partiels ont été observés.
Étant la seule espèce à présenter cette caractéristique, ce trait a évolué après la divergence évolutive de l’espèce par rapport aux autres Herichthys vivants.
La coloration noire unie est associée aux comportements compétitifs des mâles, tandis que les colorations plus claires sont associées à la formation de couples.
Cependant, d’autres variations de couleur existent, et les individus n’expriment pas toujours complètement la coloration nuptiale, il existe de nombreux changements de coloration partiels ont été observés.
Herichthys minckleyi est une espèce polygyne ce qui implique qu’un mâle se reproduit avec plusieurs femelles.
Dans ce contexte, il n’est pas rare d’observer des mâles qui gardent un territoire de plusieurs mètres carrés où se trouvent plusieurs femelles.
Pendant la période de couvage, les femelles se font discrètes jusqu’à ce que leurs alevins soient capables de nager seuls, moment auquel elles les protègent.
Après la première reproduction, il a été constaté que Herichthys minckleyi prend soin de ses jeunes différemment d’une espèce de cichlidé proche, Herichthys cyanoguttatus.
Les mâles restent moins longtemps auprès des femelles après l’accouplement et attaquent moins pour protéger leur progéniture.

Contrairement à de nombreuses caractéristiques uniques de ce poisson, cette tendance des mâles à s’absenter du nid est un trait évolutif issu de leur lignée, et non une réponse spécifique à l’espèce face aux contraintes particulières du bassin de Cuatro Ciénegas.
Herichthys minckleyi est une espèce polygyne[1], un mâle peut se reproduire avec plusieurs femelles.
De plus, les mâles gardent un territoire de plusieurs mètres carrés où se trouvent les femelles.
Pendant la période de couvage, les femelles se font discrètes jusqu’à ce que leurs alevins soient capables de nager seuls, moment auquel elles les protègent.
Après la première reproduction, il a été observé que Herichthys minckleyi prend soin de ses petits différemment d’une espèce de cichlidé apparentée, Herichthys cyanoguttatus.
Les mâles restent moins longtemps auprès des femelles après l’accouplement et attaquent moins souvent pour protéger leur progéniture.
Contrairement à de nombreuses caractéristiques uniques de ce poisson, cette tendance des mâles à s’absenter du nid est un trait évolutif issu de leur lignée et non une adaptation spécifique aux contraintes particulières du bassin de Cuatro Ciénegas.
Les valeurs de l’eau à Cuatro Ciénegas sont très stables.
Cela permet aux animaux de se reproduire toute l’année et, en outre, il ne faut pas oublier que Herichthys minckleyi est polygame.
De grands mâles sexuellement actifs occupent de vastes territoires, parfois de quelques mètres de diamètre.
Elle contient souvent plusieurs portées avec autant de femelles.
Ces nids sont constitués de grottes cylindriques excavées dans le substrat du sol.
Dans les sources plus petites, il reste souvent dans ce seul territoire.
Les tentatives de reproduction des individus les moins bien placés sont souvent la proie de la foule.
Les nids sont constitués d’environ 300 œufs qui, contrairement à de nombreux autres cichlidés méso-américains, sont pondus les uns à côté des autres.
Les œufs éclosent en moyenne au bout de deux jours et les larves restent ensuite cinq autres jours au fond du nid.
Le soin des œufs, des larves et des jeunes incombe entièrement à la femelle.
De plus, l’excavation de la grotte est généralement effectuée uniquement par la femelle.
Le mâle ne garde que son territoire, d’où la femelle bénéficie donc d’une protection indirecte.
Mais même cela n’est que de nature temporaire, car si les jeunes nagent pendant quelques jours de repos, le mâle, y compris la progéniture, est expulsé par le mâle de son territoire.
Les femelles n’ont qu’à gérer.
Ce comportement hostile aux femelles est tout à fait unique chez les cichlidés Méso-américains.
Cependant, le garçon sans père est pris en charge par la mère avec une intensité doublée et n’est jamais laissé seul.
–
La stratégie « sneakermale »
Une stratégie reproductive rare a été observée chez Herichthys minckleyi, à savoir : « La stratégie sneaker male » (OLDFIELD, 2015) : Il s’agit d’un système de reproduction de protection furtive assez répandu chez les espèces animales.
De nouvelles recherches (Université d’Otago) ont révélé comment des mâles de certaines espèces recourent à la tromperie et au déguisement pour attirer les femelles.
Il s’agit d’une stratégie dans laquelle les mâles de rang inférieur, alors que les mâles dominants luttent pour sécuriser leur territoire et accéder aux femelles, essaient de fertiliser des œufs d’un autre couple, donc ne leur appartenant pas, en effectuant une visite éclair sur le site de ponte du couple pondeur.
Il est possible que les grands territoires encouragent ce comportement et il est probable que l’élevage en grottes en soit la contre-mesure.
Il existe une lutte constante dans la nature pour améliorer le succès des accouplements.

Séquence d’événements lors de l’intrusion d’un mâle furtif. Panneau A : Un mâle furtif se cache à l’arrière-plan tandis qu’un mâle et une femelle nichent et se préparent à la ponte. Panneau B : Le mâle furtif surgit de sa cachette pour tenter une fécondation furtive. Suite à cela, le mâle nicheur mord et chasse le mâle furtif tandis que la femelle s’éloigne à la nage.
L’une des façons les plus extraordinaires dont les animaux ont répondu à ce défi est le mimétisme féminin, où les soi-disant « mâles sneaker » se déguisent en femelles pour éviter l’agressivité des mâles plus gros et voler des occasions de reproduction.
Les observateurs de ce phénomène ont été surpris de découvrir que les mâles sneakers présentaient des schémas d’expression génétique cérébrales presque identiques à ceux des femelles, mais très différents des mâles territoriaux.
Une étude a montré que les «mâles sneakers » peuvent accomplir leur exploit extraordinaire de subterfuge en activant ou désactivant des gènes spécifiques dans leur cerveau et leurs gonades.
L’étude a également révélé que les mâles sneakers se font passer pour des femelles afin de se faufiler devant d’autres mâles et d’éviter la confrontation.
L’étude a également révélé des éclairages sur la manière dont les différents mâles font face à leurs modes de vie divergents.
Les mâles sneakers expriment des gènes de neuroplasticité qui peuvent les aider à échapper aux mâles territoriaux et à voler des opportunités d’accouplement avec les femelles.
Les mâles territoriaux expriment des gènes associés au stress et à la protection contre les dommages cellulaires, ce qui suggère que la vie est difficile au sommet de la hiérarchie sociale.
Mais ce n’est pas tout ce qui se passe chez le labre à cheveux bleu qui a servi aux étude sur les mâles sneaker.
Etonnamment, les femelles peuvent changer de sexe, et les mâles sneakers peuvent changer de rôle, pour devenir des mâles territoriaux lorsqu’ils grandissent suffisamment.
L’une des façons les plus extraordinaires dont les animaux ont répondu à ce défi est le mimétisme féminin, où les soi-disant mâles sneaker se déguisent en femelles pour éviter l’agressivité des mâles plus gros et voler des occasions de reproduction.

Une étude a montré que les mâles « sneakers » peuvent accomplir leur exploit extraordinaire de subterfuge en activant ou désactivant des gènes spécifiques dans leur cerveau et leurs gonades.
Les mâles de nombreuses espèces utilisent des couleurs vives et d’autres ornementations pour attirer leurs partenaires et rivaliser avec leurs rivaux, souvent régulés par les hormones sexuelles masculines produites dans les testicules.
Souvent, il existe deux types de mâles :
- De grands mâles généralement agressifs qui courtisent ouvertement les femelles ;
- Des mâles plus petits dits « sneakers » qui ressemblent et agissent comme des femelles afin de s’accoupler en douce.
Bien qu’ils puissent ressembler et agir comme des femelles, ces mâles baskets sont parfaitement capable de se reproduire normalement puisqu’ils peuvent produire jusqu’à 60 % de spermatozoïdes en plus que les mâles territoriaux.
Une fois les œufs pondus, ceux-ci deviennent des proies pour être fécondés par les mâles sneaker qui traînent autour du nid et s’approchent pour libérer de grandes quantités de spermatozoïdes lorsqu’une femelle est en train de frayer.
Les mâles sneakers expriment des gènes de neuroplasticité qui peuvent les aider à échapper aux mâles territoriaux et à voler des opportunités d’accouplement avec les femelles.
Les mâles territoriaux expriment des gènes associés au stress et à la protection contre les dommages cellulaires, ce qui suggère que la vie est difficile au sommet de la hiérarchie sociale.
Ainsi, les petits mâles sneaker tentent de voler la fécondation tout en évitant les rencontres avec des mâles plus grands, plus agressifs et dominants.
Il est possible que les grands territoires encouragent ce comportement et il est fort probable, en retour, que l’élevage en grottes en soit la contre-mesure.
De nouvelles recherches (Université d’Otago) ont révélé comment des mâles de certaines espèces recourent à la tromperie et au déguisement pour attirer les femelles.
De l’avis de certains experts, cette stratégie de reproduction permettrait aux mâles secondaires de transmettre leurs gènes à la génération suivante, et le mécanisme de reproduction est devenu une option viable pour de nombreux mâles.
Il existe une lutte constante dans la nature pour améliorer le succès des accouplements.
Elle et ses collègues ont été surpris de découvrir que les mâles sneakers présentaient des schémas d’expression génétique cérébrales presque identiques à ceux des femelles, mais très différents des mâles territoriaux.
L’étude a également révélé que les mâles sneakers se font passer pour des femelles afin de se faufiler devant d’autres mâles et d’éviter la confrontation.
Les mâles de nombreuses espèces utilisent des couleurs vives et d’autres ornementations pour attirer leurs partenaires et rivaliser avec leurs rivaux, souvent régulés par les hormones sexuelles masculines produites dans les testicules.
Bien qu’ils puissent ressembler et agir comme des femelles, ces mâles baskets sont parfaitement capable de se reproduire normalement puisqu’ils peuvent produire jusqu’à 60 % de spermatozoïdes en plus que les mâles territoriaux.
L’étude a également révélé des éclairages sur la manière dont les différents mâles font face à leurs modes de vie divergents.
Mais ce n’est pas tout ce qui se passe chez le labre à cheveux bleu qui a servi aux étude sur les mâles sneaker.
Etonnamment, les femelles peuvent changer de sexe, et les mâles sneakers peuvent changer de rôle, pour devenir des mâles territoriaux lorsqu’ils grandissent suffisamment.
–
AVANT LA REPRODUCTION
Formation d’un couple reproducteur
Les valeurs d’eau à Cuatro Ciénegas sont très stables.
Cela permet aux animaux de se reproduire toute l’année et d’autre part Herichthys minckleyi est polygame.
Il a été constaté que plus de la moitié des couples analysés étaient issus de morphotypes différents, ou que les couples se formaient aléatoirement entre les morphotypes.
Il existe également des preuves que les mâles peuvent s’accoupler avec plusieurs femelles.
Les mâles peuvent servir simultanément plusieurs femelles pendant la période de reproduction, voire des femelles de morphes différentes.

Une femelle dans un territoire courtisera le mâle dominant, et les deux choisiront un endroit pour placer leur progéniture. L’endroit est généralement choisi sur une surface verticale près du rivage de la source ou une grotte est creusée dans les débris sous une roche, jusqu’à obtenir une surface solide.
Les grands mâles sexuellement actifs occupent de vastes territoires qui font parfois de plusieurs mètres de diamètre et dans lesquels on trouve souvent plusieurs nids avec autant de femelles.
Ces nids consistent en terriers cylindriques creusés dans le substrat du sol.
Un seul territoire d’un mâle peut contenir souvent plusieurs portées avec autant de femelles.
Dans les sources et plans d’eau de petite taille, il n’est pas rare d’observer que le mâle reste souvent dans ce seul territoire.
–
Préparation du bac
Idéalement pour reproduire cette espèce, il faudra la laisser se reproduire dans son bac de maintenance habituelle qui, éventuellement et fonction des autres espèces, sera de préférence dépeuplé des espèces de poissons présentes, principalement s’il s’agit de cichlidés.
–
Conditionnement des reproducteurs
Une alimentation principalement à base d’aliments riches en protéines aura un impact négatif sur la forme physique et la santé globale de ces cichlidés dont il faut prendre en compte les spécificités liées aux 3 types différents.
Pour autant, un apport en nourriture vivante adaptée aux besoin de chaque type aidera certainement au conditionnement et à la préparation de la reproduction.
Un complément alimentaire végétal ne devra pas être négligé.
–
Parade nuptiale
Aucune information n’est connue et n’a été rapportée à ce sujet : A COMPLETER.
–
Prémices
Herichthys minckleyi présente un fort dichromatisme sexuel en période de reproduction : Cela signifie que les mâles et les femelles sont colorés différemment, du moins lors de la phase de reproduction.
À ce moment-là, la femelle devient presque blanche tandis que le mâle devient sombre, voire presque noir.
En général, ce changement de couleurs est causé par la sélection sexuelle, mais avec Herichthys minckleyi, c’est un peu différent.
La couleur foncée des mâles dominants est principalement déclenchée par des interactions compétitives avec son propre genre.
Dans la nature, les mâles Herichthys minckleyi rivaux rôdent constamment avec l’intention permanente de prendre le contrôle du territoire et de la femelle => Cela fait référence à la stratégie reproductive rare qui a été observée chez Herichthys minckleyi, à savoir la stratégie du mâle furtif (OLDFIELD, 2015). .
C’est une stratégie dans laquelle les mâles de rang inférieur tentent de féconder les œufs lors d’une visite éclair sur le site de la ponde.
Les grands mâles sexuellement actifs occupent de vastes territoires, parfois de plusieurs mètres de diamètre et il est possible que ces grands territoires favorisent ce comportement et que la reproduction en terriers soit une contre-mesure.
Cela expliquerait aussi que les tentatives de reproduction effectuées par des individus de rang inférieur sont souvent faites au sein d’un grand groupe de ce poissons.
La couleur sombre du mâle est donc un moyen de dissuasion et en plus de cette distinction de couleur, il existe aussi une différence de longueur.
Les mâles grandissent en moyenne de 10 % de plus que les femelles et portent souvent une bosse sur le front.

Jeune mâle Herichthys minckleyi en couleurs normales.
–
Préparation du nid
Dans la nature, il semble que la femelle choisisse l’emplacement du nid, généralement une cavité offrant abri et accès restreint (KORNFIELD & Al., 1982).
L’excavation du terrier est également généralement réalisée uniquement par la femelle.

Dans un même territoire d’un mâle, il peut souvent y avoir plusieurs nids avec autant de femelles.
Ces nids creusés dans le substrat consistent en terriers cylindriques directement dans le sol.

Dans les sources plus petites, un même mâle reste souvent dans ce même territoire, pontes après pontes.
–
PENDANT LA REPRODUCTION
Ponte
Les couples pondent généralement leurs œufs dans des dépressions du fond marin ou dans de petites grottes et autres endroits offrant abri et protection.
La femelle Herichthys minckleyi choisit de préférence de pondre ses œufs sur une surface plane, parfois, il arrive que la femelle préférera pondre plusieurs centaines d’œufs sur les vitres de l’aquarium.
Plusieurs centaines d’œufs (200 à 500) ovoïdes, adhésifs et jaunâtres suivant la taille et la maturité de la femelle,, d’une longueur maximale d’environ 2 millimètres de longueur, sont déposés sur une surface préalablement nettoyée.
La femelle dépose les œufs en rangées, suivie de près par le mâle qui les féconde.
Dans certains cas, et contrairement à la plupart des cichlidés d’Amérique centrale, les œufs sont placés en grappes serrées, parfois même superposées.
Dès que la femelle a déposé ses œufs, ils sont immédiatement fécondés par le mâle.

La femelle, qui garde les œufs, retire les œufs non fécondés avec ses nageoires pectorales jusqu’à l’éclosion.
–
FECONDITE
Des données préliminaires de laboratoire indiquent une fécondité de 50 à 100 œufs pour les femelles de taille minimale (museau de 60 mm).
Les femelles dont le museau mesure 150 millimètres ont pondu entre 1 200 et 1 800 œufs en laboratoire.
–
Période d’incubation & Eclosion
La prise en charge des œufs, des larves et des petits est entièrement confiée à la femelle Herichthys minckleyi.
L’incubation dure deux jours en aquarium, et les alevins sont ensuite déposés au fond de la grotte de ponte où ils restent quatre ou cinq jours supplémentaires.
La femelle nettoie méticuleusement les œufs, retirant ceux infectés par des champignons et ceux qui sont morts, et utilise ses nageoires pectorales pour faire circuler l’eau autour d’eux.
Le mâle garde le territoire de reproduction et peut facilement intervenir en cas de danger.
La femelle défend le nid contre tous les types de prédateurs (en particulier les autres individus de la même espèce) mais c’’est elle qui s’occupe de sa progéniture.
Les œufs éclosent en moyenne après deux jours de soins et de surveillance maternels.
Les larves de poissons sont protégées par la femelle jusqu’à une taille d’environ 2 centimètres.
La femelle dépose des œufs adhésifs en rangées à la surface du nid, et le mâle passe sur la même rangée quelques secondes plus tard pour les fertiliser.
Après la ponte, la coloration de la femelle passe au blanc, tandis que celle du mâle s’assombrit.
L’éclosion a lieu en deux jours environ (selon la température), mais les larves restent fixées à la surface du nid pendant environ deux jours, période durant laquelle la femelle se déplace parfois avec ses petits.
À mesure qu’ils grandissent et commencent à se déplacer en bancs autour d’elle, elle continue de les protéger jusqu’à ce qu’ils soient capables de s’échapper seuls des prédateurs (ce qui se produit en quelques semaines, lorsqu’ils atteignent une taille de 15 à 20 millimètres).
–
Nage libre des alevins
Après l’éclosion, les larves restent au fond du nid pendant encore cinq jours, temps nécessaire pour résorber leur sac vitellin et commencer à nager.

–
Garde parentale
Le couple défend son territoire autour du nid avant et après la ponte, mais le mâle abandonne généralement la femelle peu après.
Le mâle ne garde que son territoire, dont la femelle bénéficie donc d’une protection indirecte.
Mais cette protection passive n’est que temporaire, car si les jeunes nagent librement pendant quelques jours, la femelle, y compris les petits, est expulsée de son territoire par le mâle.
Les femelles Herichthys minckleyi doivent alors se débrouiller seules…avec leurs couvées !
La femelle solitaire reste près de ses alevins, les guidant et leur communiquant par des mouvements spasmodiques de ses nageoires.
Elle ne les quitte jamais.
Les alevins se nourrissent apparemment de détritus.
Quelques semaines plus tard, lorsqu’ils atteignent une taille suffisante (environ 2 centimètres) pour explorer seuls, ils commencent à désobéir à leur mère et finissent par chercher refuge dans les zones peu profondes et végétalisées où ils passent leur vie juvénile.

Ce comportement misogyne est complètement unique chez les Cichlidés méso-américains.
Cependant, les jeunes orphelins de père sont soignés par la mère avec une intensité redoublée et ne sont pas laissés seuls un instant.
Le mâle ne garde que son territoire, dont la femelle bénéficie donc d’une protection indirecte.

La femelle protège les œufs et les alevins pendant plusieurs semaines, jusqu’à ce qu’ils atteignent un stade de développement suffisant pour échapper à la plupart des prédateurs.

Suffisamment grands, ils rejoignent alors les berges et les zones peu profondes offrant un couvert suffisant pour se protéger et grandir seuls.
A ce stade, la femelle retrouve sa coloration normale.

Les femelles Herichthys minckleyi guident les alevins sur le fond marin grâce à des mouvements saccadés de leurs nageoires et de leur corps, tandis que les alevins se nourrissent de détritus organiques et de petites particules comestibles qu’ils trouvent sur leur chemin.

https://www.facebook.com/pronaturane/videos/2040938882856039/
Lorsqu’ils atteignent une taille suffisante pour se débrouiller seuls, environ 20 millimètres de longueur, ils commencent à quitter leur mère et à se réfugier sous la végétation surplombant les sources.

C’est à partir de ces zones qui leur offrent une certaine sécurité qu’ils s’aventurent progressivement au cours de leur croissance à partir en quête de nourriture jusqu’au moment où ils auront atteint une taille adulte qui leur permettra d’évoluer sereinement.
–
Première alimentation des alevins
Comme les autres espèces d’Herichthys, les alevins sont assez faciles à élever.

En bac de croissance, ils acceptent les nauplies d’artémias et autres aliments fins dimensionnés à la taille de leur bouche.
Plus que tous les autres alevins de cichlidés d’Amérique centrale, ils ont besoin constamment d’eau propre et dépourvue de toute pollution chimique car lorsqu’ils sont dans un bac surpeuplé où les conditions de pureté de l’eau se dégraderont rapidement, ils peuvent très vite être sujets à développer des ballonnements du ventre, qui chez les alevins sont fondamentalement incurables.
Par conséquent, il faudra éviter de vouloir éviter d’élever trop d’alevins dans un aquarium trop petit.
–
CONSERVATION
GENERALITES
En tant qu’espèce en danger, le cichlidé de Minckley fait l’objet de travaux de conservation et une gestion éclairée est cruciale.
Les espèces endémiques sont particulièrement vulnérables à la destruction de leur habitat, ce qui ajoute davantage de défis de conservation.
De plus, comme Herichthys minckleyi présente plusieurs morphes, les poissons sont répartis dans différents habitats du bassin de Cuatro Ciénegas.
L’implication conservatoire de ce polymorphisme est de protéger l’hétérogénéité des conditions du substrat, afin que tous les morphes puissent avoir leur habitat nécessaire.

Actuellement, l’aire de répartition de cette espèce endémique est protégée par le gouvernement fédéral mexicain, classée comme « Area de Protección de Flora y Fauna » et gérée par la « Secretaría del Medio Ambiente y Recursos Naturales ».
Le bassin de Cuatro Ciénegas est également un site de conservation d’intérêt pour le Fonds mondial pour la nature, l’UNESCO et la Nature Conservancy.
Espérons que les protections accordées au bassin pourront également contribuer à la conservation et à la récupération du cichlidé unique de Minckley.
En tant qu’espèce unique, endémique et en danger, le cichlidé de Minckley a attiré des chercheurs du monde entier pour comprendre le polymorphisme trophique.
De plus, ce poisson apporte une contribution précieuse à la biodiversité dans une région du monde incroyablement unique et unique.
–
Suivi des populations
L’Aire Protégée (APFFCC) considère cette espèce comme une cible prioritaire de suivi.
Un projet de marquage-recapture (utilisant des puces PIT) mené par l’auteur dans un bassin (Mojarral Oeste = Poza Azul) en est à sa sixième année et a permis de suivre plus de 2 000 individus marqués.
Bien que les résultats n’aient pas encore été publiés, le programme fournit des données très utiles pour les études et le suivi de la biologie des populations, ainsi que des informations sur les déplacements des poissons entre les différents sites.
–
Tendances démographiques
Il n’existe aucune estimation publiée du nombre total d’individus. Des estimations préliminaires, basées sur un seul échantillon (données non publiées de HENDRICKSON), indiquent des fluctuations annuelles, mais aucune conclusion ne peut être tirée quant aux tendances à long terme.
Cependant, il ne fait aucun doute que la population totale de l’espèce a diminué au cours des 50 dernières années, en raison de l’assèchement de plusieurs habitats qu’elle occupait auparavant (notamment la rivière Garabatal).
–
INTRODUCTION EN DEHORS SON BIOTOPE
Sans objet ⇒ Herichthys Minckleyi nécessite des conditions de maintenance particulière qui n’autorisent pas une diffusion de cette espèce de cichlidé.
Tout le commerce semble provenir d’élevages en captivité et non de captures directes à Cuatro Ciénegas ; il ne représente donc pas une menace directe pour les populations sauvages.
–
ACTIONS DE CONSERVATION
En tant qu’espèce menacée, Herichthys Minckleyi fait l’objet d’efforts de conservation et une gestion éclairée est essentielle. Les espèces endémiques sont particulièrement vulnérables à la destruction de leur habitat, ce qui complexifie encore leur conservation.
De plus, comme Herichthys Minckleyi présente plusieurs morphotypes, les poissons sont répartis dans différents habitats du bassin de Cuatro Ciénegas.
Ce polymorphisme a pour enjeu la conservation de l’hétérogénéité des substrats, afin que chaque morphotype puisse bénéficier de l’habitat nécessaire.
Actuellement, l’aire de répartition de cette espèce endémique est protégée par le gouvernement fédéral mexicain, classée « Aire de protection de la flore et de la faune », et gérée par le Secrétariat à l’environnement et aux ressources naturelles.
Le bassin de Cuatro Ciénegas est également un site d’intérêt pour la conservation, reconnu par le WWF, l’UNESCO et The Nature Conservancy.
On espère que les mesures de protection accordées au bassin contribueront également à la conservation et au rétablissement d’Herichthys Minckleyi.
–
Menaces
Catégories et facteurs de risque
La modification des habitats constitue sans aucun doute la menace la plus importante, notamment la déviation des cours d’eau et l’assèchement des zones humides, qui entraînent des variations drastiques du niveau et de la température de l’eau.
L’agriculture intensive à grande échelle, utilisant l’irrigation par pompage des eaux souterraines des vallées adjacentes, représente une menace sérieuse.
Si la connexion entre le système hydrologique souterrain de la Valle del Hundido (au sud de la vallée de Cuatro Ciénegas) et celle de la vallée de Cuatro Ciénegas fait encore l’objet de nombreux débats parmi les hydrologues, il n’en va pas de même pour la connexion entre la Valle Calaveras (au nord) et Cuatro Ciénegas.
Le pompage des eaux souterraines dans la Valle Calaveras a rapidement épuisé la ressource en eau alimentant la commune de Cuatro Ciénegas, avec de graves conséquences pour cette dernière.
La source et la zone de recharge des aquifères alimentant les bassins de la vallée étant encore inconnues, aucune mesure de contrôle n’a été mise en œuvre.
Il est donc essentiel et urgent de mener des études hydrologiques fournissant des informations précises sur l’origine et le volume de la nappe phréatique, ainsi que sur sa zone et son niveau de recharge.
Les plantes invasives représentent un risque considérable, notamment les espèces « Arundo donax » et « Tamarix ramosissima ».

Arundo donax.

Tamarix ramosissima.
Très consommatrices d’eau, elles peuvent accroître l’évapotranspiration et, par conséquent, abaisser le niveau et le débit des mares et des rivières.
De plus, elles risquent de modifier la géomorphologie des cours d’eau et des mares et de limiter l’ensoleillement, pourtant crucial pour les écosystèmes de la vallée.
Le tourisme a déjà un impact significatif, en particulier pendant la Semaine sainte, lorsque des milliers de visiteurs affluent aux mares de La Becerra et de Churince, sur la rivière Mezquites et à Las Playitas, malgré leur situation en zone protégée.
Un suivi rigoureux est donc nécessaire dans toute la vallée, et l’accès à de nombreuses zones essentielles à la conservation des espèces doit être interdit.
Le tourisme a également entraîné l’introduction de nouvelles espèces exotiques et favorisé leur dispersion vers de nouveaux habitats.
Ces espèces représentent une menace importante, probablement accrue par d’autres impacts physiques et chimiques.
Dans certaines régions, la contamination par les déchets humains et les ordures ménagères constitue une menace supplémentaire.
L’espèce est principalement menacée par l’assèchement dû à l’exploitation agricole de la nappe phréatique, entraînant la destruction de certains points d’eau.
On observe également un dessèchement, une déforestation et une modification des plans d’eau à des fins touristiques, ainsi que la construction de canaux d’irrigation et de routes.
Le surpâturage par les chevaux et les chèvres, ainsi que le brûlage des pâturages pour le bétail, constituent d’autres menaces.
L’exploitation des ressources se poursuit, mettant en péril les poissons, les mollusques, les crustacés et les reptiles.
L’aquaculture à Cuatro Cienegas d’espèces commerciales telles que :
- Ictalurus lupus ;
- Micropterus salmoides (Achigan à grande bouche).
sont des menaces (ARRIAGA-CABRERA & Al., 1998) pour :
- Cichlasoma cyanoguttatum ;
- Herichthys minckleyi ;
- Lepomis megalotis.
De façon générale, le drainage et les espèces exotiques menacent la vallée.
Beaucoup de ces biotopes désertiques isolés, en particulier dans le sud des États-Unis, ont disparu au fil du temps.
Les conséquences dans un tel cas sont bien plus graves que, par exemple, l’assèchement d’un bras fluvial.
Dans ce dernier cas, il y a tout simplement moins d’habitat pour une espèce, mais dans le premier, un ensemble génétique est éradiqué.
L’eau est souvent extraite pour produire de l’alimentation animale.
La libération d’espèces exotiques représente également une menace croissante.
Par exemple, Herichthys guttatus (le cichlidé rouge d’Afrique) se trouve en grand nombre.
Le gouvernement fédéral a tenté d’éradiquer cette espèce, mais a dû abandonner ces tentatives en raison de coûts excessifs et d’un résultat décourageant.
En 1994, Cuatro Ciénegas a été déclaré réserve naturelle protégée.
–
Mesures de conservation
Présente dans l’aire protégée de Cuatro Ciénagas, l’espèce ne fait l’objet d’aucune mesure de conservation connue.
Elle est inscrite à la NORMA OFICIAL MEXICANA : NOM-059-SEMARNAT-2010 comme espèce endémique et menacée.
–
USAGES HUMAINS
Aquariophilie : Très limitée en raison de la difficulté à maintenir cette espèce de cichlidé en aquarium.
La maintenance de ce poisson, hors normes habituelles, s’adresse de préférence à des spécialistes de l’aquariophile bien équipés.
–
MARCHE AQUARIOPHILE
Quasi nul à très peu.
L’espèce est difficile à maintenir en aquarium et peu populaire auprès des aquariophiles, hormis les plus passionnés.
L’espèce est rarement commercialisée, mais a été observée dans quelques animaleries aux États-Unis et en Europe, apparemment issue d’élevages en captivité (observation personnelle à Austin, Texas, États-Unis).
–
MENACE POUR LES HUMAINS
Sans objet.
Ces poissons sont inoffensifs.
–
STATUT DE CONSERVATION
Statut IUCN
Actuellement, Herichthys Minckleyi est classé comme « ESPECE EN DANGER » sur la Liste rouge de l’UICN, ce qui signifie qu’il existe un risque élevé d’extinction de ce poisson à l’état sauvage.

Mise à jour du 19 Octobre 2018
Des informations supplémentaires sont nécessaires concernant la taille et l’évolution de la population de cette espèce, mais on sait qu’elle est en déclin là où son habitat a été desséché en raison de la surexploitation des ressources en eau (D. HENDRICKSON, 2018).
Référence : 4836
–
FishBase
https://www.fishbase.se/summary/Herichthys_minckleyi.html
–
CITES
Non évalué.
–
AUTRES DONNEES
Wikiespèces : Herichthys minckleyi
Cichlasoma minckleyi
–
REFERENCES
KORNFIELD I. & TAYLOR J. 1983. Une nouvelle espèce de poisson polymorphe Cichlasoma minckleyi, de Cuatro Ciénegas, Mexique
HENDRICKSON, D. ; MEJIA GUERRERO, O. ; PEREZ‐MIRANDA, F. (2019). « Herichthys minckleyi ». Liste rouge UICN des espèces menacées. 2019 e.T4836A3012480. doi :10.2305/UICN. UK.2019-2.RLTS.T4836A3012480.en. Consulté le 16 novembre 2021.
KORNFIELD, Irv ; TAYLOR, JEFFERY N. (1983). « Une nouvelle espèce de poisson polymorphe, Cichlasoma minckleyi, originaire de Cuatro Ciénegas, Mexique (Teleostei : Cichlidae) ». Actes de la Société biologique de Washington. 96 : 253–269.
OLDFIELD, RONALD G. ; MANDREKAR, KAPIL ; NIEVES, M. XAVIER ; HENDRICKSON, DEAN A. ; CHAKRABARTY, PROSANTA ; Swanson, Brook O. ; Hofmann, Hans A. (01-04-2015). « Soins parentaux chez le cichlidé de Cuatro Ciénegas, Herichthys minckleyi (Teleostei : Cichlidae) ». Hydrobiologie. 748 (1) : 233–257. Bibcode : 2015HyBio.748.. 233O. doi :10.1007/s10750-014-2081-4. ISSN 1573-5117.
PEREZ-MIRANDA, FABIAN ; MEJIA, OMAR ; SOTO-GALERA, EDUARDO ; ESPINOSA-PEREZ, HECTOR ; PIALEK, LUBOMIR ; ŘICAN, OLDRICH (2018). « Phylogénie et diversité des espèces du genre Herichthys (Teleostei : Cichlidae) ». Journal of Zoological Systematics and Evolutionary Research. 56 (2) : 223–247. doi :10.1111/jzs.12197. ISSN 1439-0469.
MAZA-BENIGNOS, Mauricio De La (2014), SISTEMÁTICA, ZOOGEOGRAFÍA Y EVOLUCIÓN DEL GÉNERO HERICHTHYS (PISCES : CICHLIDAE), EN EL NORESTE DE MÉXICO, Inédit, doi :10.13140/2.1.2764.7047, consulté le 13-11-2025
« Ordre CICHLIFORMES : Famille CICHLIDAE : Sous-famille CICHLINAE (d-w) ». Le projet ETYFish. 22-08-2018. Consulté le 13-11-2025.
CHAKRABARTY, PROSANTA (2004). « Biogéographie des ciclidés : commentaire et critique ». Poissons et Pêcheries. 5 (2) : 97–119. Bibcode : 2004FiFi…. 5…97C. doi :10.1111/j.1467-2979.2004.00148.x. ISSN 1467-2979.
PEREZ-MIRANDA, FABIAN ; MEJIA, OMAR ; LOPEZ, BENJAMIN ; ŘICAN, OLDRICH (29-04-2020). « Horloges moléculaires, biogéographie et diversité des espèces à Herichthys avec évaluation du rôle de Punta del Morro comme frein vicaire le long de la zone de transition mexicaine dans le contexte de la diversification locale et globale des cichlidés ». PeerJ. 8 e8818. doi :10.7717/peerj.8818. ISSN 2167-8359. PMC 7195834. PMID 32391194.
SOUZA, VALERIA ; SIEFERT, JANET L. ; ESCALANTE, ANA E. ; ELSER, JAMES J. ; EGUIARTE, LUIS E. (2012). « Le bassin des Cuatro Ciénegas à Coahuila, Mexique : un parc astrobiologique précambrien ». Astrobiologie. 12 (7) : 641–647. Bibcode : 2012AsBio.. 12..641S. doi :10.1089/ast.2011.0675. ISSN 1531-1074. PMC 3426885. PMID 22920514.
MINCKLEY, W. L. (1984). « Cuatro Cienegas Fishes : Revue de la recherche et test local de diversité versus taille d’habitat ». Revue de l’Académie des Sciences de l’Arizona-Nevada. 19 (1) : 13–21. ISSN 0193-8509. JSTOR 40024286.
HULSEY, C. D. ; MARKS, J. ; HENDRICKSON, D. A. ; WILLIAMSON, C. A. ; COHEN, A. E. ; STEPHENS, M. J. (2006). « Spécialisation alimentaire chez Herichthys minckleyi : un poisson polymorphe trophiquement ». Journal of Fish Biology. 68 (5) : 1399–1410. Bibcode :2006JFBio.. 68.1399Herichthys doi :10.1111/j.0022-1112.2006.01021.x. ISSN 1095-8649.
KORNFIELD, IRV ; SMITH, DAVID C. ; GAGNON, P. S. ; TAYLOR, JEFFREY N. (1982). « Le poisson ciclidé de Cuatro Ciénegas, Mexique : preuve directe de la conspécificité entre morphes trophiques distincts ». L’évolution. 36 (4) : 658–664. doi :10.2307/2407880. ISSN 0014-3820. JSTOR 2407880.
HULSEY, C. DARRIN ; GARCIA-DE-LEON, FRANCISCO J. (2013). « Hybridation introgressive chez un cichlidé trophiquement polymorphe ». Écologie et évolution. 3 (13) : 4536–4547. Bibcode : 2013EcoEv… 3.4536Herichthys doi :10.1002/ece3.841. ISSN 2045-7758. PMC 3856752. PMID 24340193.
SKULASON, SKULI ; SMITH, THOMAS B. (1995-09-01). « Polymorphismes de ressources chez les vertébrés ». Tendances en écologie et évolution. 10 (9) : 366–370. Bibcode : 1995TEcoE.. 10..366S. doi :10.1016/S0169-5347(00)89135-1. ISSN 0169-5347. PMID 21237070.
HULSEY, CRISTOPHER ; EHEMANN, NICOLAS (juin 2025). « Cuatro Ciénegas, las « Islas Galápagos » de Coahuila : un modèle para étudier la évolution de cíclidos mexicanos » (PDF). SIMAC Boletín. 59 : 10–14.
HULSEY, C ; HENRICKSON, D ; DE LEON, F (2005). « Morphologie trophique, performance alimentaire et utilisation des proies chez le poisson polymorphe Herichthys minckleyi ». Recherche en écologie évolutive. 7 (2) : 303–324.
SWANSON, BROOK O. ; GIBB, ALICE C. ; MARKS, JANE C. ; HENDRICKSON, DEAN A. (2003). « Le polymorphisme trophique et les différences comportementales diminuent la compétition intraspécifique chez un cichlidé, Herichthys minckleyi ». Écologie. 84 (6) : 1441–1446. Bibcode : 2003Ecol… 84.1441S. doi :10.1890/02-0353. ISSN 1939-9170.
KOCHER, THOMAS D. (2004). « Évolution adaptative et spéciation explosive : le modèle du poisson cichlidé ». Nature critique Génétique. 5 (4) : 288–298. doi :10.1038/NRG1316. ISSN 1471-0064.
« La Liste rouge des espèces menacées de l’UICN ». Liste rouge UICN des espèces menacées. Archivé depuis l’original le 24-09-2025. Consulté le 13-11-2025.
SYKES, L ; SANTINI, L ; ETARD, A ; NEWBOLD, T (2020). « Effets de la forme de rareté sur les réponses des espèces à l’utilisation des terres ». Biologie de la conservation. 35 (3) : 688–696. Bibcode : 2020ConBi… 34..688S. doi :10.1111/cobi.13419. PMID 31532012.
COHEN, ADAM E. ; HENDRICKSON, DEAN A. ; PARMESAN, CAMILLE ; MARKS, JANE C. (2005). « Ségrégation de l’habitat parmi les morphes trophiques du Cichlidé Cuatro Ciénegas (Herichthys minckleyi) ». Hydrobiologique. 15 (2) : 169–181. ISSN 0188-8897.
HENDRICKSON, D. ; MEJIA GUERRERO, O. ; PEREZ‐MIRANDA, F. (2019). « Herichthys minckleyi ». Liste rouge UICN des espèces menacées. 2019 e.T4836A3012480. doi :10.2305/UICN. UK.2019-2.RLTS.T4836A3012480.en. Consulté le 16 novembre 2021.
KORNFIELD, IRV ; TAYLOR, JEFFERY N. (1983). « Une nouvelle espèce de poisson polymorphe, Cichlasoma minckleyi, originaire de Cuatro Ciénegas, Mexique (Teleostei : Cichlidae) ». Actes de la Société biologique de Washington. 96 : 253–269.
OLDFIELD, RONALD G. ; MANDREKAR, KAPIL ; NIEVES, M. XAVIER ; HENDRICKSON, DEAN A. ; CHAKRABARTY, PROSANTA ; SWANSON, BROOK O. ; HOFMANN, HANS A. (01-04-2015). « Soins parentaux chez le cichlidé de Cuatro Ciénegas, Herichthys minckleyi (Teleostei : Cichlidae) ». Hydrobiologie. 748 (1) : 233–257. Bibcode : 2015HyBio.748.. 233O. doi :10.1007/s10750-014-2081-4. ISSN 1573-5117.
PEREZ-MIRANDA, FABIAN ; MEJIA, OMAR ; SOTO-GALERA, EDUARDO ; ESPINOSA-PEREZ, HECTOR ; PIALEK, LUBOMIR ; ŘICAN, OLDRICH (2018). « Phylogénie et diversité des espèces du genre Herichthys (Teleostei : Cichlidae) ». Journal of Zoological Systematics and Evolutionary Research. 56 (2) : 223–247. doi :10.1111/jzs.12197. ISSN 1439-0469.
MAZA-BENIGNOS, MAURICIO DE LA (2014), SISTEMÁTICA, ZOOGEOGRAFÍA Y EVOLUCIÓN DEL GÉNERO HERICHTHYS (PISCES : CICHLIDAE), EN EL NORESTE DE MÉXICO, Inédit, doi :10.13140/2.1.2764.7047, consulté le 13-11-2025
« Ordre CICHLIFORMES : Famille CICHLIDAE : Sous-famille CICHLINAE (d-w) ». Le projet ETYFish. 22-08-2018. Consulté le 13-11-2025.
CHAKRABARTY, PROSANTA (2004). « Biogéographie des ciclidés : commentaire et critique ». Poissons et Pêcheries. 5 (2) : 97–119. Bibcode : 2004FiFi…. 5…97C. doi :10.1111/j.1467-2979.2004.00148.x. ISSN 1467-2979.
PEREZ-MIRANDA, FABIAN ; MEJIA, OMAR ; LOPEZ, BENJAMIN ; ŘICAN, OLDRICH (29-04-2020). « Horloges moléculaires, biogéographie et diversité des espèces à Herichthys avec évaluation du rôle de Punta del Morro comme frein vicaire le long de la zone de transition mexicaine dans le contexte de la diversification locale et globale des cichlidés ». PeerJ. 8 e8818. doi :10.7717/peerj.8818. ISSN 2167-8359. PMC 7195834. PMID 32391194.
SOUZA, VALERIA ; SIEFERT, JANET L. ; ESCALANTE, ANA E. ; ELSER, JAMES J. ; EGUIARTE, LUIS E. (2012). « Le bassin des Cuatro Ciénegas à Coahuila, Mexique : un parc astrobiologique précambrien ». Astrobiologie. 12 (7) : 641–647. Bibcode : 2012AsBio.. 12..641S. doi :10.1089/ast.2011.0675. ISSN 1531-1074. PMC 3426885. PMID 22920514.
MINCKLEY, W. L. (1984). « Cuatro Cienegas Fishes : Revue de la recherche et test local de diversité versus taille d’habitat ». Revue de l’Académie des Sciences de l’Arizona-Nevada. 19 (1) : 13–21. ISSN 0193-8509. JSTOR 40024286.
HULSEY, C. D. ; MARKS, J. ; HENDRICKSON, D. A. ; WILLIAMSON, C. A. ; COHEN, A. E. ; STEPHENS, M. J. (2006). « Spécialisation alimentaire chez Herichthys minckleyi : un poisson polymorphe trophiquement ». Journal of Fish Biology. 68 (5) : 1399–1410. Bibcode :2006JFBio.. 68.1399H. doi :10.1111/j.0022-1112.2006.01021.x. ISSN 1095-8649.
KORNFIELD, IRV ; SMITH, DAVID C. ; GAGNON, P. S. ; TAYLOR, JEFFREY N. (1982). « Le poisson ciclidé de Cuatro Ciénegas, Mexique : preuve directe de la conspécificité entre morphes trophiques distincts ». L’évolution. 36 (4) : 658–664. doi :10.2307/2407880. ISSN 0014-3820. JSTOR 2407880.
HULSEY, C. DARRIN ; GARCIA-DE-LEON, FRANCISCO J. (2013). « Hybridation introgressive chez un cichlidé trophiquement polymorphe ». Écologie et évolution. 3 (13) : 4536–4547. Bibcode : 2013EcoEv… 3.4536H. doi :10.1002/ece3.841. ISSN 2045-7758. PMC 3856752. PMID 24340193.
SKULASON, SKULI ; SMITH, THOMAS B. (1995-09-01). « Polymorphismes de ressources chez les vertébrés ». Tendances en écologie et évolution. 10 (9) : 366–370. Bibcode : 1995TEcoE.. 10..366S. doi :10.1016/S0169-5347(00)89135-1. ISSN 0169-5347. PMID 21237070.
HULSEY, CRISTOPHER ; EHEMANN, NICOLAS (juin 2025). « Cuatro Ciénegas, las « Islas Galápagos » de Coahuila : un modèle para étudier la évolution de cíclidos mexicanos » (PDF). SIMAC Boletín. 59 : 10–14.
HULSEY, C ; HENRICKSON, D ; DE LEON, F (2005). « Morphologie trophique, performance alimentaire et utilisation des proies chez le poisson polymorphe Herichthys minckleyi ». Recherche en écologie évolutive. 7 (2) : 303–324.
SWANSON, BROOK O. ; GIBB, ALICE C. ; MARKS, JANE C. ; HENDRICKSON, DEAN A. (2003). « Le polymorphisme trophique et les différences comportementales diminuent la compétition intraspécifique chez un cichlidé, Herichthys minckleyi ». Écologie. 84 (6) : 1441–1446. Bibcode : 2003Ecol… 84.1441S. doi :10.1890/02-0353. ISSN 1939-9170.
KOCHER, THOMAS D. (2004). « Évolution adaptative et spéciation explosive : le modèle du poisson cichlidé ». Nature critique Génétique. 5 (4) : 288–298. doi :10.1038/NRG1316. ISSN 1471-0064.
« La Liste rouge des espèces menacées de l’UICN ». Liste rouge UICN des espèces menacées. Archivé depuis l’original le 24-09-2025. Consulté le 13-11-2025.
SYKES, L ; SANTINI, L ; ETARD, A ; NEWBOLD, T (2020). « Effets de la forme de rareté sur les réponses des espèces à l’utilisation des terres ». Biologie de la conservation. 35 (3) : 688–696. Bibcode : 2020ConBi… 34..688S. doi :10.1111/cobi.13419. PMID 31532012.
COHEN, ADAM E. ; HENDRICKSON, DEAN A. ; PARMESAN, CAMILLE ; MARKS, JANE C. (2005). « Ségrégation de l’habitat parmi les morphes trophiques du Cichlidé Cuatro Ciénegas (Herichthys minckleyi) ». Hydrobiologique. 15 (2) : 169–181. ISSN 0188-8897.
–
LITTERATURE
- ARTIGAS AZAS JM 1996. Les cichlidés du désert
- COHEN AE &Al. 2005. Ségrégation de l’habitat parmi les morphes trophiques des cichlidés de Cuatro Ciénegas (Herichthys minckleyi)
- HULSEY CD &Al. 2006. Spécialisation alimentaire à Herichthys minckleyi, un poisson polymorphique trophique
- HULSEY CD &Al. 2015. Dimorphisme sexuel chez un poisson cichlidé polymorphique trophique
- KORNFIELD I. & TAYLOR J. 1983. Une nouvelle espèce de poisson polymorphe Cichlasoma minckleyi, de Cuatro Ciénegas, Mexique
- MINCKLEY WL 1969. Environnements du Bolson de Cuatro Cienegas Coahuila au Mexique, avec une référence spéciale au biote aquatique
- OLDFIELD RG &Al. 2015. Soins parentaux dans le cichlidé Cuatro Cienegas, Herichthys minckleyi
- RIBBENS P. 2008. Cichlidés dans le désert, Herichthys minckleyi (KORNFIELD &TAYLOR, 1983) Cichlidae 34 – 6, déc. 2008.
- SWANSON BO & Al., 2003. Le polymorphisme trophique et les différences de comportement diminuent la compétition intraspécifique chez un cichlidé, Herichthys minckleyi
–
LEXIQUE
[1] Le travertin est une roche sédimentaire calcaire continentale à aspect concrétionné, grossièrement litée, de couleur blanche quand elle est pure, ou tirant vers le gris, le jaunâtre, le rougeâtre ou le brun, selon les impuretés qu’elle renferme.
La roche est caractérisée par de petites cavités (vacuoles) inégalement réparties.
Cette roche carbonatée se dépose aux émergences de certaines sources, et dans des cours d’eau peu profonds à petites cascades.
Les travertins actuels contiennent de l’aragonite qui recristallise rapidement en calcite.
Selon Julien Curie, le mot travertin désigne la roche produite en condition thermale (eau naturellement réchauffée), alors que le tuf calcaire désigne la roche produite dans une eau froide.
[2] Hyalella est un genre de crustacés amphipodes de la famille des Hyalellidae.
[3] Mexithauma quadripaludium est une espèce d’escargot d’eau douce, un mollusque gastéropode aquatique de la famille des Hydrobiidae.
Cette espèce est endémique de la vallée de Cuatro Ciénegas, dans le désert de Chihuahuan, au Mexique.
[4] Un stromatolithe ou stromatolite (on parle aussi parfois de « thrombolite », est une structure laminaire constituée de calcaire, parfois de silice, qui se développe en milieu aquatique peu profond, marin ou d’eau douce continentale.
Les stromatolithes sont d’origine à la fois biogénique (biolithogenèse par des communautés de cyanobactéries) et sédimentaire (structures carbonatées en feuillets empilés formés de particules sédimentaires piégées dans des voiles algaires gélatineux produits par ces bactéries).
[5] Le prognathisme (du latin pro, « avant » et du grec « γνάθος », « mâchoire ») est une configuration faciale selon laquelle une ou les deux mâchoires sont projetées en avant par rapport à la « verticale » allant du front au nez.
[6] La polygynie, où un mâle s’accouple avec plusieurs femelles tandis que chaque femelle s’accouple avec un seul mâle, est considérée comme le système d’accouplement fondamental des animaux.

Vous devez être connecté pour poster un commentaire.